


1 孤独的天才
为坚持智力追求,不惜放弃其天伦之乐;
在学术群体外围,做出科学的核心发现;
用数学分析生物,成功地进行学科交叉;
十年一系列实验,一篇论文开创新学科。
他孤立于当时的科学界,做出奠基性突破却终生未被学界承认;他的工作几十年后尚不为同一学科的诺贝尔奖得主所理解;他发现的貌似简单的理论,大多数学过其结论的人,都没意识到其智力高度;他不是为利益做研究的纯粹科学家,身后却被疑造假,遭遇不公。
这位孤独的天才,就是自称为“实验物理学教师”的遗传学之父:孟德尔(Johann Gregor Mendel,1822-1884)。
我认为,生物学有两座智力高峰:第一次是1854年至1866年孟德尔独自一人;第二次是1951年至1965年克里克(Francis Crick)及其合作者们。两个高峰碰巧相隔一百年。
今天重读孟德尔的论文,追寻孟德尔的思路,思考孟德尔的环境,仍然很有意义。
1.1 孟德尔的论文和思路
由于同时代理解孟德尔科学工作重要性的人极少,他的遗物保留下来的很少。孟德尔最重要的遗物是一篇遗传学论文。与此相关,他还有一篇遗传学论文以及给一位植物学家的10封信。
他的主要论文显示了清晰的思路,有助于我们追踪科学是如何在一个头脑中诞生的。
孟德尔的时代,人们对遗传的认识还很粗浅,基本认同“混合遗传”(blending inheritance)学说:遗传是“黑+白=灰”,父母的黑和白简单融合得到子代的灰。此学说虽未被正式提出和论证,却是一个普遍接受的、朴素的、以为不证自明的规律。
而孟德尔不以为然,他设计实验,通过锲而不舍的研究,发现了与此不同的学说。从1854年开始,孟德尔用豌豆做了一系列遗传学实验,时间长达十年。他于1865年公布所发现的遗传学规律,并于次年以德文在《布鲁恩自然史学会杂志》发表了论文《植物杂交的实验》(Versuche über die Pflanzen-Hybriden)。
从孟德尔的文章,我们可以体会他如何做研究:发现重要问题,提出解决问题的途径;设计实验思路,进行实验研究;得到结果,分析结果,提出前人没有想到的理论;进一步实验,得到更多可以分析的结果;进而推广理论, 证明理论。
孟德尔的论文由十一部分组成。
在“引言”部分,孟德尔简要回顾以往研究后,立即明确提出问题:无人成功地提出过对杂交体形成和发生普遍适用的规律。
他指出前人做过不少杂交实验,但未得到普遍规律是因为所需的工作不仅量大,而且较难。孟德尔认为需要考虑到:规模要相当大;具有不同型的杂交后代要定量分析;在不同代间要准确地知道不同型之间的关系;要确切地分析它们之间的相对数量关系。
他写道:需要勇气花力气做大量实验,这是唯一正确的道路,才能最终解决重要的问题。..本文就是仔细研究的结果,进行了八年的工作得出的结论。
孟德尔说的八年,是他收集论文所用数据的八年。其实,此前,他做了两年实验,摸索条件选择最佳材料。所以实际上,论文工作进行了十年。十年实验后,又隔两年才发表论文。论文发表后,他还用其他植物做过几年研究。
在“实验植物选择”部分,孟德尔指出:“任何实验的价值和用处取决于所用材料是否符合其目的,所以选什么植物和怎么做实验并非不重要…必须特别小心地选择植物,从开始就避免获得有疑问的结果。”
他选的植物首先具有恒定的分化特征;其次,在进行杂交的时候不会受到外来花粉的污染;另外,每一代杂交后代生殖力不能变。
孟德尔所谓“分化特征”现在称为“性状”(如高矮、颜色);他的“恒定”是指同一性状在不同代之间不变;他注意避免外来花粉污染,怕不确切知道父本,研究结果无从分析;他还注意代间生殖力无变化,减少在性状数量分析时的干扰。
后人认为,为了选择到合适的实验材料,孟德尔有可能考虑过二十多种植物。孟德尔说他因为花形状的奇异而试了豆科(Leguminosae),后决定用豌豆(Pisum)。对所用豌豆的确切生物学分类,孟德尔并不是很确定,说“专家意见”说大多数是Pisum sativum,还有几种,不过他明确指出分类对其研究并不重要。
用豌豆还有论文中没说明的、实验操作的优点:既能自花授粉,又能异花授粉,较易人为控制。1854年和1855年,孟德尔试过34种不同的豌豆。在孟德尔为数不多的遗物中,有一张1856年购买豌豆的订单。
在“实验分工和安排”中,孟德尔对所研究的性状进行了选择:他选择成对的性状,研究他们在代间的传递规律。这些性状可以在代间稳定遗传,且易于识别和区分。
在“杂交体的外形”部分,他进一步说明了对性状的选用。他专门选择子代性状一定相同于父本或母本的性状,而不是介于父母之间、或其他变异。孟德尔知道豌豆有些性状居于父母母本之间,而不等同于父本、或母本,例如,在论文第八部分,他发现杂交体的开花时间介于父母本之间,但孟德尔没有研究介于父母本性状之间的性状。他研究的7对性状,一定是与父本或母本相同,每对中必定有一种传到下代,而一对性状的两种在后代不会变化,也不会永远消失。孟德尔明确这样选择的重要性。
孟德尔的选择简化了分析从而可以得出有意义的结论。比如到2017年知道,有八百多个基因决定人的高矮,子代高矮是父母这些基因及其含有的更多多态性综合结果,另外还有环境因素(如食物)等。如果谁在十九世纪研究人身高的遗传,就很难得出简单的规律,这并非人类高矮不符合孟德尔遗传规律,而是很难进行分析。
他选了7对性状:种子形状(平滑或皱褶)、种子颜色(黄或绿)、种皮颜色(紫或白)、豆荚形状(鼓或狭)、豆荚颜色(紫或白)、花的位置(顶或侧)、茎的高度(长或短)。
对应于7对性状,孟德尔安排了7个实验。实验一用15株植物做了60次授粉;实验二用10株植物做了58次授粉;实验三用10株植物做了35次授粉;实验四用10株植物做了40次授粉;实验五用5株植物做了23次授粉;实验六用10株植物做了34次授粉;实验七用10株植物做了37次授粉。
所有实验,孟德尔都进行了双向杂交:一对性状中,如种子颜色的黄和绿,既做过父本黄、母本绿,也做过父本绿、母本黄,他发现亲本来源不影响这些性状的传代。
他认识到性状有显隐之分,发明了“显性”(dominant)和“隐性”(recessive)两个词。当父本母本分别是不同性状(如黄和绿),而他们杂交子代只显现一种性状(黄)时,孟德尔称显现的一种(黄)为显性、没有显现的(绿)为隐性。他指出,隐性在杂交体(以后称为F1代)看不见,但在以后可以完全不变地重新显现。进一步的实验表明:显性隐性与父本母本来源也无关。
他确定了7对性状的显隐性:种子形状平滑为显而皱褶为隐、种子颜色黄为显而绿为隐、豆荚颜色黄为显而绿为隐、豆荚形状鼓为显而狭为隐、花色紫为显而白为隐、花的位置顶为显而侧为隐、茎的高度长为显而短为隐。“任何实验中都没有过渡型式”。
我们现在知道,其实在两年的预实验中,孟德尔实际上得到了纯合子。虽然当时并无纯合子和杂合子的概念,他本人也未明确这样说,但如果不以纯合子开始实验,分析结果会很复杂。
在孟德尔所谓“杂交体来的第一代”实验结果部分,我们稍需改变他的称呼,以方便叙述。他用的第一代,我们现在称为F0代。他所谓“杂交体”,我们现在称F1代。他称“杂交体来的第一代”,现称F2代。
我们可以看到,他用不同表型的两种F0代亲本间授粉得到的F1代均表现显性的性状,比如,豌豆种子分别为平滑和皱褶的F0代父母本授粉得到的F1代的豌豆种子都是平滑的,没有皱褶的。
接着,他让F1代自花授粉,得到F2代,发现隐性(如皱褶)没有因为在F1代不表现而永远消失,它重新出现在F2代。进一步数量分析表明,在F2代,显性对隐性呈3:1的比例。孟德尔强调,3:1比例毫无例外地适用于所有(7对)性状。其中,实验一发现:从253株F1代杂交体得到7324颗F2代种子,其中5474颗平滑,1850颗皱褶,比例为2.96:1。实验二发现:258株F1代植物产生了8023颗F2代种子,其中6022颗种子黄色,2001颗绿色,比例为3.01:1。
孟德尔还分析每个豆荚内种子形状和颜色是否有关,不同植物是否有关,结果认为都无关。他指出如果算的植物少了,比例漂移很大;如果昆虫损害了种子,也会影响对性状的确定。
从实验三到实验七,他列出了其他5对性状的传代结果,发现7对性状平均显隐比例为2.98:1。他看到了规律:F1代100%为显性;F2代隐性重现,而且有规律,显隐比例3:1。
孟德尔知道隐性没有在F1代不表现而消失,所以知道混合学说不对。至此,他已经超出一般人,而他还继续迈出了后面三步,发现3:1的比例、探究对比例的解释、用实验验证解释,从而获得了新的理解,远远超过了同时代的其他生物学家,包括所有时代最伟大的科学家之一。
孟德尔在看到3:1的比例后,他分析在F2代显性的性状可以有两种意义,它可以是F0一样(自交后保持与亲代的显性性状),或是F1代(自交后既有显性性状的后代、也有隐性性状的后代)。只能用F2代再做一代实验来检验是哪种状况。他预计,如果F2和F0一样,那么其后代性状就应该不变,而如果F2代类似F1杂交体状态,那么其行为与F1相同。
由此,引出孟德尔下一年的实验,即他所谓“杂合体来的第二代”(我们现称F3代)部分结果。他发现,表现隐性性状的F2代,传F3代后其性状不再变化(总是隐性表型)。而表现显性的F2代,其F3代结果表明:2/3的F2代是杂交体(其F3代出现3:1的显性和隐性),而另外1/3的F2代其F3代都是显性表型。
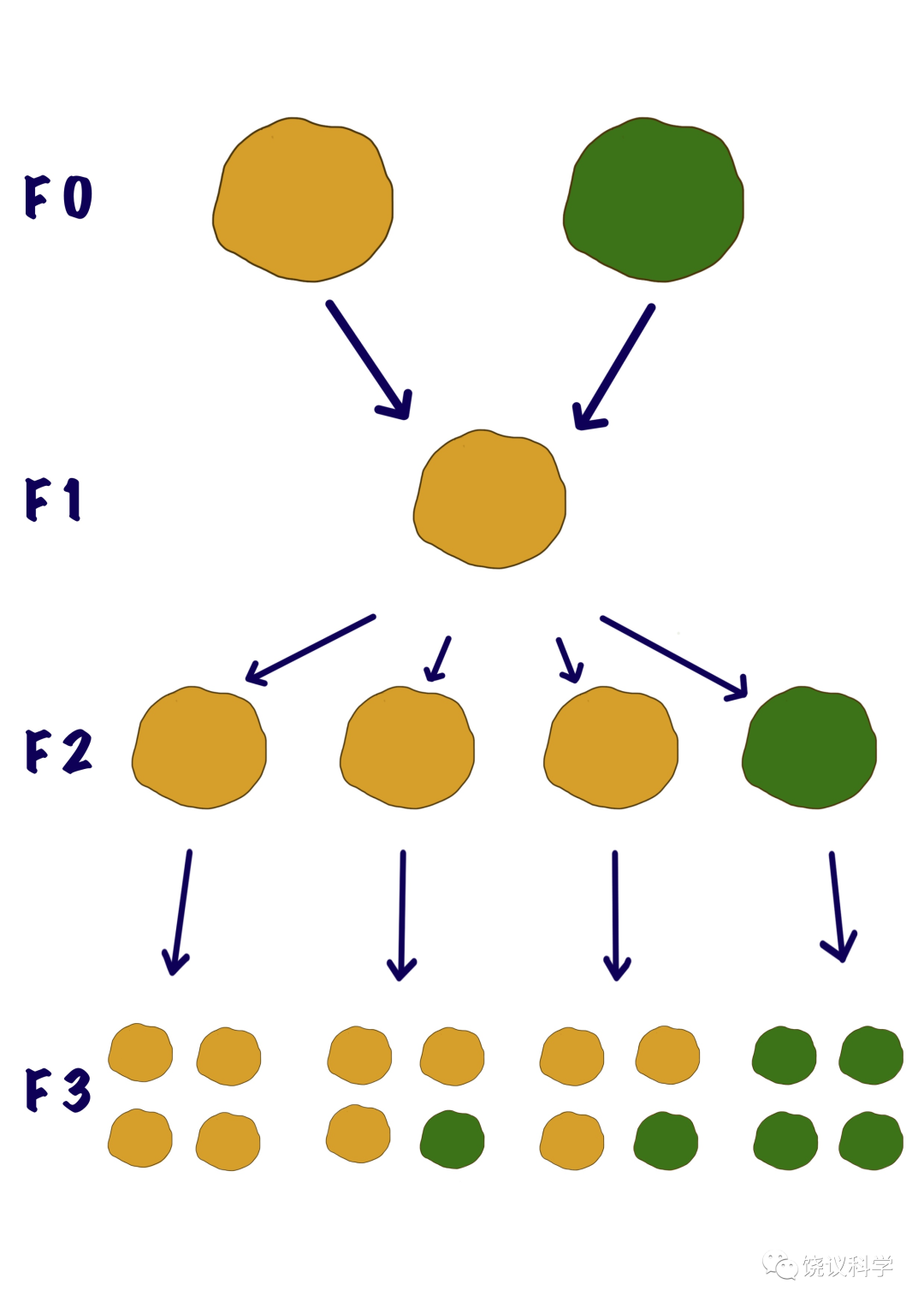
实验一:F2代565棵平滑豌豆植株,193株F2代只产生平滑的F3种子,372株F2代生平滑和皱褶的F3种子(F3的平:皱比例为3:1)。也就是说,F2代中显性的其实含类似F0和类似F1的比例为1.93:1。
实验二:F2代519棵黄色种子的豌豆植株自交后,166株只生黄色种子的F3代,353株生黄和绿种子(黄:绿为3:1),F2代中表现显性的植株含类似F0和类似F1的比例为2.13:1。
从实验三到实验七算其他五种性状时,孟德尔没有每次都算全部后代性状,而只分析100株植物的后代,结果有漂移但大体相似。他说计算数量大的实验一和实验二更有意义。实验五漂移最大,他重复了一次,数字更趋接近预计比例。
这样,孟德尔将F2的3:1中的3,进一步分成2和1。3:1就被分解成1:2:1(显性恒定:杂交体:隐性恒定)。
在F3代后,他还做了几代“杂交体后几代”,发现结果都符合F3代前所发现的规律,“没有察觉任何偏移”。到发表论文时,实验一和二做了六代,实验三和七做了五代,实验四、五、六做了六代。可以算出,他用豌豆做了17610次授粉。
孟德尔再迈进了一步:数学模型。
生物学研究用数学的较少。即使是今天,虽然有些生物学家非常需要定量,但绝大多数生物学研究者关心数量只在乎升高、降低和不变。孟德尔以数量分析、定量不同表型的植物,从而发现3:1的规律,继而推出和验证1:2:1的规律,已经使他成为在生物学领域成功运用数学的先驱。
在以上基础上,孟德尔进一步用了数学模型。这就超出不仅那时、甚至包括今天绝大多数生物学研究者。他提出,用A表示恒定的显性,a表示恒定的隐性,Aa表示杂合体。那么F2代就是:A+2Aa+a。
他观察到的F2代1:2:1就符合这个数量关系(杂合性状为2,显性和隐性恒定性状皆为1)。
分别分析单个性状传代情况后,孟德尔研究了不同对的性状间是否有关系。在“几个分化性状相关联杂交体的后代”部分,孟德尔发现7对性状之间完全独立。比如种子是平滑还是皱褶,与种子是黄色还是绿色毫无关联。总结这部分实验结果,孟德尔说:每对不同性状之间的关系独立于亲本其他不同(性状)。
后人好奇,为什么孟德尔做的7对性状都无关?如果有些基因在染色体较近位置的话,会有一定关联。现在知道,他做的7对性状,其基因分别在4条染色体上,而在同一染色体上的三个正好分别在染色体上相距很远的位置。
孟德尔在发现各对性状独立传代后,他在文章中可能考虑了自己的发现与进化论的关系。我们现在知道,他读过第二版《物种起源》德译本,在书的边缘做了评注。可能由于自己在修道院吃饭,他不能公开说接受进化论,所以在论文中完全没提进化论。但是,他文章故意讨论了性状独立遗传的意义。他指出:如果一个植物有7种不同的性状,产出后代就有2的7次方(128)种不同的组合。孟德尔的这个算法其实解决了 “混合学说”给达尔文进化论造成的矛盾。混合学说导致每一代比上一代更少样(黑加白得到灰,灰加灰得此灰,以此类推),而不是多样,可供选择的越来越少,生物应该退化。而孟德尔推出不同组合的数量很多,每代的多样性在增加,进化就有很多可以选择。
行文至此,孟德尔简要总结了结果:分化性状在杂交组合中行为完全一模一样。每对分化性状杂交体的后代,一半又是杂交体,另外一半中含同等比例的亲本恒定分化性状。(这等于是他用文字复述1:2:1的发现)。如果不同分化性状在杂交时组合起来,每对分化性状成为组合系列。
孟德尔也认为通过研究他选择的性状所得到的规律,也适用于其他的性状。
在从外观的性状上推出规律后,孟德尔推断外观的差别实际是由生殖细胞的组成的差别所造成。原因在于雄性的花粉细胞,雌性的卵细胞。
他推理:因为总是当卵细胞和花粉细胞具有同样的恒定性状(显性纯合子或隐性纯合子)时,其后代得到同样的恒定性状(显性或隐性纯合子),所以此时两种细胞都有创造同样个体的物质。我们必须认为在杂交体(显性和隐性杂合子)授粉后出现恒定性状(显性或隐性纯合子)时,也是这样。“杂交体的卵巢中卵细胞的种类,或杂交体雄蕊中花粉细胞数量,与可能的恒定组合型式相同,卵细胞和花粉细胞的内在组分与其不同外形相符”(斜体为本文所加)。
如果F0代是恒定的显性,其生殖细胞应该内含A的花粉细胞和内含A的花粉细胞。如果F0代表型是恒定隐性,其生殖细胞应该内含a的花粉细胞和内含a的卵细胞。F0代花粉细胞和卵细胞交配后,得到F1代。F1代的花粉细胞有A和a两种、且数量相等,卵细胞也有数量相等的A和a两种。在F1代自交时,各自含A或a的两类花粉细胞与各自含A或a的卵细胞交配后,不同花粉细胞有同等机会与不同的卵细胞组合,那么得到的下代就有:A/A, A/a, a/A, a/a等四种。其中Aa和aA个体不同只在于其显性隐性来源不同,一个来源卵细胞,一个来源花粉细胞,但最后表型相同,可以归为Aa。这样,F2就应该是A+2Aa+a。

F1代产生F2代可以表示为:
A/A+A/a+a/A+a/a= A+2Aa+a。
孟德尔这个等式很重要。他将等式左边性细胞内的成分和右边得到植物后代的表型连起来。左边是我们现在说的基因型,右边是表型。孟德尔从表型的1:2:1推导出生殖细胞遗传物质的组成。他依据的是观察到的表型,推测生殖细胞的情形。
孟德尔说明这是平均的结果,具体每个后代有多种可能,而且随机,所以分开的实验肯定有漂移,只有大量收集数据,才能得到真实的比例。在这里,我们可以猜想孟德尔意识到了纯合子A/A,a/a和杂合子A/a 和a/A,可惜没有明确提出名词。
至此,他把理论深入到生殖细胞,而且可以用数学模型表示遗传学的规律,虽然其数学虽然简单,是很基本的组合。数学分析结合生物学实验,产生很重要的意义,揭示了遗传的规律。
因为孟德尔希望找到普遍适用的规律,所以,他论文最后一部分实验是“其他种属植物杂交体的实验”,检验他从豌豆发现的规律是否适用于其他植物。在论文发表时,他说开始用了几种其他植物,其中用大豆做的两个实验已经做完。用Phaseolus vulgaris和Phaseolus nanus(两者都是菜豆)做的杂交结果, 发现后代好几个性状的传代完全吻合符合他从豌豆得到的规律。但是,用Ph nanus和Ph multiflorus做杂交时,其花色有较多变异。孟德尔觉得花色仍符合他发现的遗传规律,提出要假设花色是两个或更多独立颜色的组合,花色A由单个性状A1+A2+…..的组合而成。他实际上提出了多基因遗传,而通常误解导致“孟德尔遗传学”被误认为单基因遗传学。
孟德尔经过新颖的、严谨的、长期的实验和定量分析,终于找到了杂交发育的普适规律。后人将孟德尔发现的规律表述成为两个定律:第一个是分离律,决定同一性状的成对遗传因子彼此分离,遗传给后代,也可以表述为颗粒遗传,以区别于以前流行的混合学说,说明因子没有消失;第二个是自由组合律,确定不同遗传性状的遗传因子间可以自由组合(本章省略了孟德尔原文研究不同性状A、B、C之间的关系部分)。虽然这些内容在原文中都有叙述,孟德尔本人并不认为自己发现了两个分开的规律,而是一个普遍的规律。
在“结语”部分,孟德尔介绍前人杂交实验的结果和前人有关植物受精过程的论述。他指出:根据著名生理学家的意见,植物繁殖时,一个花粉细胞和一个卵细胞结合成为单个细胞,同化和形成多个新细胞,长成植物个体。
然后孟德尔提出:(杂交体)发育遵循一个恒定的定律,其基础就是细胞中生动地结合的“因子的物质组分和安排(material composition and arrangement of elements)”。…豌豆的胚胎毫无疑问是亲本两种生殖细胞中因子的结合。…如果生殖细胞是同类的,那么新个体就像亲本植物…如果杂交后代不同,必需假设卵细胞和花粉细胞的分化因子间出现妥协,形成作为杂交体基础的细胞,但矛盾因子的安排只是暂时的,…分化的因子在生殖细胞形成时可以自我解放。在生殖细胞形成时,所有存在的因子完全自由和平等地参与,分化的因子互相排斥地分开。这样,产生卵细胞和花粉细胞的种类在数量上相同于形成因子可能的组合数量。
将孟德尔原文的“因子”换成现代的“基因”,就可以几乎原封不动地以他的文字理解遗传。对于喜欢直观的人来说,还有一个总结孟德尔的简单方法是:A/A+A/a+a/A+a/a。
孟德尔文中六次复述相似的内容:豌豆杂交形成生发细胞和花粉细胞,其中的组成数量相同于通过授粉将性状组合起来的所有恒定型式。这也表明他知道遗传的基础在于生殖细胞中存在数量相应于性状的物质。
在1870年9月27日,孟德尔给植物学家Nägeli的信中明确用anlage(德文“原基”)描述遗传因子,也说明他对基因的理解与现在很接近。
孟德尔早年研究过老鼠毛发颜色的遗传,被要求停止:修道院不宜做动物交配。他自己做道长后,1871年在花园建蜂房,用蜜蜂做过实验,但未见报道蜜蜂遗传结果,所以没有将植物中发现的规律推广到动物。
1.2 其他科学家对遗传学的理解
孟德尔时代的科学家如何理解遗传?孟德尔时代的科学家如何理解孟德尔?孟德尔之后第二伟大遗传学家如何理解孟德尔?我们可以讨论三位科学家:孟德尔同代的Nägeli、达尔文和四十年后的摩尔根。
孟德尔寄出40份论文单行本给不同科学家,其中,只有瑞士著名植物学家、慕尼黑大学教授Nägeli回了信。所以,40人中Nägeli算最重视孟德尔。孟德尔不仅给Nägeli寄了论文,而且他们还交换了植物种子。孟德尔自己提出用山柳菊做实验验证豌豆中发现的规律,得到研究山柳菊的专家Nägeli的鼓励。孟德尔信中说过种子少、不容易授粉、自己时间少。1867年11月6日他给Nägeli的信还说“老天让我过度肥胖,使我不再适合做植物园户外工作”。 他得到结果有点慢,不知情的会以为他在找借口、磨洋工。等他把山柳菊实验做完后,发现不符合豌豆里面得出的规律。孟德尔在信中告诉Nägeli,山柳菊的结果和豌豆的矛盾,但自己还做了其他植物,紫罗兰、茯苓、玉米和紫茉莉,发现结论和豌豆一样,所以山柳菊比较特殊,而自己发现的规律适用于多数植物。Nägeli不为所动,尽管孟德尔写过很多信告诉他辛辛苦苦做的实验,Nägeli发表植物学重要著作时,一字不提孟德尔的工作。正确地解释山柳菊结果要等到1904年,山柳菊是单性繁殖(所谓孤雌生殖),所以不能父本母本杂交,而遗传规律其实和豌豆相同。
仅以Nägeli的例子,还不能说孟德尔是超越时代的天才,而比较达尔文更说明问题。
1859年,达尔文发表《物种起源》提出了进化论,其核心是:“如果出现对生物生存有利的变异, 有此特性的个体就一定会有最佳的机会在生存斗争中保存下来;这些个体在强大的遗传原理中倾向于产生有类似特性的下一代。我把这一保存原理,或适者生存,称为自然选择。”如何遗传是达尔文自然选择进化论的必要支柱,达尔文非常希望了解遗传学。
神学对达尔文的攻击虽然猛烈,但非理性。而有人提出了严厉而富有逻辑的理性批评:自然选择进化论违背当时人们理解的遗传规律共识。根据“混合学说”,生物的性状黑加白得到后代灰,灰加灰出现的后代次灰,依此类推,性状越来越单调,不存在很多可供选择的性状,因此没有物竞天择的物质基础。所以,达尔文急需遗传学说为进化论提供解释和支持。但是,遗传规律在他眼皮底下溜过去了。
与一般人印象不同,达尔文不仅依赖观察来推导理论,他也做过实验。达尔文用花做了十一年的实验,部分结果先于孟德尔于1862年以论文形式发表,主要结果发表于1876年和1877年的两本书中,也散在于其他书中。
1868年,达尔文发表《动植物在家养情况下的变异》。此书记录了达尔文用金鱼草做的实验。常见金鱼草的花是双侧对称(达尔文称common型式,我们表为大写C),但偶尔也会出现一些怪怪的金鱼草变种,其花呈现辐射对称(达尔文称peloric型式,我们表为小写p)。达尔文把具有p性状的父本与具有C性状母本进行杂交,发现所得后代(F1代)全部呈现C性状。进一步授粉得到127株F2代金鱼草中,88株具有C性状,37株具有p性状,2株介于两种性状之间。他的实验到此结束。
观察到实验结果后,达尔文的结论是:同种植物里有两种相反的潜在倾向,…第一代是正常的占主要,…隔一代怪的倾向增加。
这样的结论没有太大意义,远不如孟德尔深刻,即使不做实验的人们也能通过生活经验得到直观的“常识”。
达尔文不止一次失去机会。在1877年的《同种植物不同花型》一书中,从他总结的报春花研究结果的表格中,我们可以看到,他用杂合体授粉时,得到显性后代为75%,隐性为25%,一个完美的3:1。不过,达尔文还是没有意识到其重要性,再次与现代遗传学失之交臂。
在《动植物在家养情况下的变异》中,达尔文提出了错误的泛生论(pangenesis)。他提出生物体全身体细胞都产生泛子gemmules(后人亦称pangenes),进入性细胞中,这些gemmules组合决定了性细胞内含,形成不同的性细胞,再产生不同的后代。在强调体细胞产生泛子的重要性时,达尔文说生殖能力要么不全在于生殖细胞,要么生殖细胞没有生殖能力,而是收集和选择泛子。他论述此假说时,将代间遗传、植物嫁接、发育、再生等多种现象混在一起谈,认为有同样机理。他的讨论相当于混淆了我们现在知道的细胞全能性(很多细胞本身含有整套遗传物质)、与代间遗传两个不同层次的问题。他在讨论中接受拉马克主义的“用进废退”,而认为泛生假说能解释用进废退,受外界影响的体细胞性状可以获得并通过gemmules进入性细胞而传代。现代科学表明,生物体中无泛子。后人从pangenesis这个词中抽出了gene来表示基因。
对比孟德尔的实验和推理,可以看到达尔文的问题:1)达尔文没有意识到样本量太小,实验设计有问题,没有做到孟德尔论文很前面就提到的“从开始就避免获得有疑问的结果”;2)达尔文在获得F1代的结果看到都是C性状时,和其他做杂交实验观察到同样现象的人一样,没有提出显性和隐性的概念;3)F2代重新出现F1代不见了的p性状,达尔文也仅看到现象,提出所谓“回复原理”(Principle of Reversion)复述现象,并无原理;4)在F2得到数量时,他没算两种性状的比例(2.38:1),也不知道比例蕴含的意义;5)没有推测而发现下一步的1:2:1;6)没有数学模型;7)没有从实验结果中发现规律,提出错误的遗传理论。
我们不知道达尔文是否读过孟德尔的文章。有些人认为,假如达尔文读了,也读不懂,或者不能接受孟德尔的理论。我们知道孟德尔在达尔文1860年第二版《物种起源》的德译本上有批注。孟德尔1866年的论文有时好像是他希望给达尔文的进化论提供遗传基础。孟德尔从自己发现的多个性状自由组合规律,推算如果有7对不同性状的两种植物间授粉,可以产生很多不同的组合,从而解释了多样性。孟德尔很可能在1866年就想到了自己发现的规律对于进化论的意义。当然,孟德尔当时的实验没有考虑进化论还需要的一部分:变异如何出现。要等七十年后,到1930年代后,英国的费舍尔(Ronald A Fisher,1890-1962)和霍尔丹(JBS Haldane,1892-1964)、美国的莱特(Sewall Wright,1889-1988)和杜布赞斯基(Theodosius Dobzhansky,1900-1975)等才成功地将孟德尔遗传学和达尔文进化论结合起来。
一般教科书说三位科学家1900年重新发现孟德尔:德国的Carl Correns (1864-1933)、荷兰的Hugo de Vries (1848-1935) 和奥地利的Erich von Tschermak (1871-1962),虽然von Tschermak已被遗传史学家排除在重新发现者之外。这几位所谓重新发现孟德尔的人,理解程度当时都还低于孟德尔。de Vries重新写数学公式不如35年前孟德尔的公式。三人的工作量加起来也远不如孟德尔一人。Correns是Nägeli的学生和亲戚,推动了对孟德尔的认识。英国的William Bateson (1861-1926) 对孟德尔学说的推广起了很大作用。
第二伟大的遗传学家,无疑是美国的摩尔根(Thomas H. Morgan,1866-1945)。但是,直到1909年,摩尔根还发表文章称孟德尔的方法是玩数字的高级杂耍(superior jugglery)。事实上,摩尔根当年不仅不信孟德尔,也不信遗传的染色体学说。是1910年他自己发现了白眼突变果蝇的事实后,他也做了和孟德尔一样的交配实验,取得数据和比例。为了解释事实,摩尔根不得不沿着孟德尔的思路,也提出因子,也进行拼凑数字的“高级杂耍”,最后奠定了遗传学的现代基础。在事实面前,摩尔根不得不“出尔反尔”,因为科学真理高于个人偏见,也不会败于俏皮话的讥笑挖苦。
Nägeli的狭隘、达尔文的缺憾、摩尔根的态度,给孟德尔的超前程度提供了绝佳的注释。
1.3 孟德尔的生平
孟德尔出生地德文称Heinzendorf,捷克称Hyncice,现在捷克境内,当时属于奥匈帝国。孟德尔的父亲是佃农,每周四天料理自家的田地,三天给一位女伯爵干农活。命运似乎注定了孟德尔不得不子承父业,终其一生在农田中度过,但当地的神父Johann A.E. Schreiber (1769-1850)鼓励孟德尔的父母让他多受教育。孟德尔自己也要与命运抗争,并得到姐妹的支持。孟德尔后来为报答妹妹的支持,资助了她的孩子读书。
1850年4月17日,他为了考教师证以第三人称写过一个自我简介,清楚地说明了他的情况、心境和决心,信的大意是:
…小学后,1834年他上中学。4年后,接连不断的灾难[译注:一次是他父亲事故受伤],使他父母完全不能支持他学业所需的费用。因此,16岁的他落入不得不完全自己支持自己的可悲境地。所以,他一边给人做家教,一边上学。1840年中学毕业时,首要问题是取得必要的生活来源。因此,他曾多次试图做家庭教师,由于没有朋友和推荐,未果。失去希望和焦虑的痛苦、未来前景的悲观,彼时对他有强烈影响,导致生病,被迫和父母待了一年。次年,他努力后得以做私人教师,以支持学业。通过极大努力后,他成功地修完两年的哲学。他意识到无法这样继续下去,所以在学完哲学后,他觉得非得进入一个生命驿站,能让自己脱离痛苦的生存挣扎。他的境况决定了他的职业选择。
1843年,他要求并得以进入布鲁诺的圣汤玛斯修道院。从此,他的物质境况彻底改变。有物质生活的舒适后,他重新获得勇气和力量。他满心欢喜和集中精力学习经典。空余时间忙于修道院一个小型植物和矿物收藏。有机会接触后,他对自然科学的特别爱好更加深化。…虽然缺乏口头教育,而且当时教学方法特别困难,从此他却更依附于自然研究。他努力通过自学和接受有经验者的教诲,来弥补自己的缺陷。1845年,他到布鲁诺哲学学院听了农业、园艺和葡萄种植课程。…他很乐意代课,倾力以容易理解的方式教学生,并非无成效…
孟德尔坦陈入修道院不是为了宗教信仰,而是经济原因。这一重要的人生选择中他权衡的不是神圣与世俗,而是智力追求与成家育子的权利。为了头脑,他舍弃了生殖权。对于血气方刚的青年,并非容易,而需要很大的决心。孟德尔的决定也和中国传统的一种说法(也是当代相当一部分华人的想法)不同:这些人读书是为了颜如玉,而孟德尔为了智力追求放弃颜如玉。
1843年,不满21岁的孟德尔进入布鲁恩(Brünn,现称Brno)的圣汤玛斯修道院(the Abbey of St. Thomas),并于1847年25岁成为神父。孟德尔原名Johann,入修道院后加Gregor教名。
到修道院后,他同时做过代课老师。那时,中学老师已需要证书。孟德尔第一次教师资格考试没通过,被送到维也纳大学去学习,这加强了他的科学背景。孟德尔曾再考教师资格,还是没能通过,而且,估计两次都是没过生物学,所以后来只能做代课老师,在当地的实科中学(Brünn Realschule)教了14年低年级物理学和自然史。他一直以实验物理学教师自称,而不说是生物学家。
孟德尔积极参与学术活动。他长期研究气象,曾任国家气象和地磁研究所布鲁恩站长,1862年提交布鲁恩地区15年气象总结。他一生中参与了八个科学学会、二十六个非科学协会。1861年,孟德尔在任课的中学和一百多人共同创立当地的自然史学会。1865年2月8号和3月8号两个星期三的晚上,在布鲁恩自然科学学会,孟德尔宣读了豌豆研究结果。当地小报对孟德尔演讲有报道,但未能引起国际科学界的注意。
1866年论文发表后,孟德尔将40份抽印本寄给国际上的科学家,后人找到了13份的下落,传说达尔文处有,并未证实。发表文章的杂志有120本在世界主要图书馆。
1868年,修道院道长去世后,孟德尔经过两轮选举后当选道长。他不用教书后,但还有其他工作繁重,他还是尽量做了研究。他用了多种植物做遗传实验。留下的纸片表明在去世前三年,他还在想有关豌豆的遗传问题。1865年到1878年,他记录了14年的地下水位。1870年,他加入养蜂协会,1877年报告对蜜蜂飞行和产蜜量的四年观察。他曾研究苹果和梨的抗病性。在一些协会刊物中,他以M和GM笔名写过一些短篇。
孟德尔生活丰富。他的政治观点偏自由派,与自己的教会背景矛盾。而他支持的自由派掌政时,出台的税收政策却对他的修道院很不利。政府为缓和与他争论曾安排他任银行副董事长和董事长。但他持续十年坚决反对税收,造成他晚年生活很大的苦恼。他在政治上左右碰壁。
1884年1月6日,孟德尔去世。他生前要求尸检,结果表明他肾炎并发心脏病。有位年轻的神父将其诗化,称孟德尔是心给伤了。孟德尔自己是乐天派,年纪大的时候回顾自己一生满意多于不满意。
园艺协会刊物讣告称:“他的植物杂交实验开创了新时代”。 猜想讣告作者是刊物主编Josef Auspitz(1812-1889),他曾任实科中学校长,支持孟德尔无证代课14年,是孟德尔的重要支持者和欣赏者之一。 但是,讣告的溢美之辞远非共识。
据他的朋友Gustav von Niessl (1839-1919)说,孟德尔生前相信“我的时代会到来”。确实如此。但是,要等他去世16年、理论公布34年以后。
1900年声称重新发现孟德尔的三位科学家。其中de Vries的第一篇论文没有提孟德尔,后来可能因为隐瞒不住曾借鉴孟德尔的事实(包括难以解释如果他没有读过孟德尔,为什么他第一篇文章用了孟德尔的dominant和recessive两个词),在第二篇论文中说是重新发现孟德尔。von Tschermak可能不懂孟德尔也说自己重新发现了孟德尔,所以史学家认为不能算。有趣的是,von Tschermak的外公 Eduard Fenzl(1808-1879)是维也纳大学教孟德尔的生物老师之一,不仅教学保守,也可能是没让孟德尔第二次考到教师证书的考官之一。
1.4 孟德尔“造假”案
除了有人说孟德尔不懂自己发现了什么以外,对于孟德尔最大的冤枉是说他编造了实验结果。英国统计学家和遗传学家费舍尔于1936年首先发难,他对孟德尔的实验数据进行统计分析后,断定孟德尔的数据过于接近理想数据。轻一点说,孟德尔可能有我们不知道的助手,在做了前两年实验导致孟德尔有理论后,助手为了满足孟德尔的理论而在后面几年给孟德尔提供他喜欢的数据。重一点说就很难听:“多数—如果不是所有—的实验结果都伪造了,以期贴切地符合孟德尔的预期”。以后每过一些年,就有人小聪明又发现孟德尔的“问题”。
反击孟德尔造假说法的文章也不断。最近一篇较好的反击是2007年哈佛大学Hartl和Fairbanks 发表于《遗传》杂志的文章。
我认为,给孟德尔伸冤的首要理由是:他无需造假。科学对于他来说不能带来利益。他如果造假,最对不起的是放弃生育人权、十几年如一日做研究的他自己。
其次,孟德尔时代没有统计学。统计学是几十年以后发明的。孟德尔只需分析数量关系,无需检验统计显著性。那时不知道应该做多少次实验、收集多少数据后才应该停止实验。可能是孟德尔收集到觉得差不多的就时候停止,所以数据会接近预计。孟德尔也在论文中明确说过,有一次实验漂移较远,他重复了实验后,数据更接近预计。
孟德尔的行为证明他不是造假和隐瞒不利结果的人。他曾努力使怀疑自己工作重要性的Nägeli相信自己发现的规律。但即使这种情况下,他也没隐瞒自己发现了有悖于自己理论的现象。他把自己的豌豆种子给了Nägeli和其他人,希望他们验证自己的结果。孟德尔致Nägeli信说:我观察到山柳菊的杂交行为与豌豆的正好相反。孟德尔用另外四种植物(紫罗兰、茯苓、玉米和紫茉莉)做的实验观察到其杂交行为都与豌豆一样。
孟德尔不仅在给Nägeli的信说明了山柳菊的结果,而且将结果在1869年发表了。后来多年认为,有两种遗传方式,一种是“豌豆式”(符合经典孟德尔学说),一种是“山柳菊式”(不符合孟德尔学说)。虽然以后也发现这些生物其实都符合孟德尔学说,造成困惑是因为山柳菊是单性遗传,但当时孟德尔以为山柳菊与豌豆不同。如果孟德尔造假,或选择只符合自己理论的结果,那么他就无需在已经公开自己的理论后,将只有他自己知道的山柳菊的结果直接告诉一位不愿接受自己理论的人,而且发表第二篇生物学论文,公布与第一篇的矛盾。
1.5 孟德尔的精神遗产
孟德尔以天生的才能、青年的果断和壮年的坚持,在困难中成长,以放弃获得条件,在失败中得机遇,最终在有限的环境做出了超越时代的发现。
孟德尔的成就,一百多年来催生了多个现代科学学科。首先是直接导致遗传学诞生,而对于同时期诞生的进化论,孟德尔可能隐约知道自己工作的意义,虽然遗传学和进化论结合于1930年代。二十世纪遗传学与生物化学结合,并与微生物、生物物理学交叉,在1940年代又催生了分子生物学。1970年代诞生的重组DNA技术,全面改观了生命科学:分子生物学深入到从医学到农业各个领域,带来多个学科的变革,人类遗传学、基因组学、生物信息学是其直接传承。
在应用上,遗传学带来了二十世纪绿色革命,对于解决全人类食物起了很大作用。遗传学、分子生物学和重组DNA技术奠定了现代生物技术、产生了生物技术产业。现代遗传学和基因组学为个体化医学奠定了必不可少的基础,虽然我们今天还远未达到个体化医学的远景。
孟德尔的发现,对于科学和人类,今后长期还将有深远影响。
最后的问题是:既然孟德尔不受科学家重视,不为科学界所认同,那么,他怎么能获得做研究的条件?
这个问题,背后有一个更加鲜为人知的故事:欲知后事如何,请听下回分解……
注1:孟德尔用“杂交”一词,是现代意义的cross(动物可译成“交配”、植物“授粉”),而非后来科学家重新定义的“杂交”,即 不同种或不同品系之间的交配。孟德尔文章中多半都是同种植物的交配,并非物种或品系间的交配。“杂交”一词今天在中国学生和老师中仍未严格使用,部分原因可能是学孟德尔理论时听惯了杂交一词。
注2:本文中斜体都是孟德尔原文的着重强调。
注3:孟德尔的论文中用了“对照实验”(control)一词。每个在野外做的实验,他都在暖房中也做了,证明野外实验未因昆虫或外源花粉等环境因素所干扰,结果可信,他才采用。
注4:孟德尔用花粉细胞来表示精细胞。现在知道花粉中包含2或3个细胞。参与受精的是其中的两个精细胞。
注5:孟德尔在结语中说花粉细胞和卵细胞结合成单个细胞后,“同化和形成多个新细胞”。现在看来“同化”是错误的,限于当时对发育的误解。全部细胞都来源于受精卵分裂、增值,并不发生同化母体细胞参与子代发育。
参考文献
http://www.mendelweb.org/
Corcos A and Monaghan F (1985) Role of de Vries in the recovery of Mendel's work. I. Was de Vries really an independent discoverer of Mendel?Journal of Heredity 76:187-90.
Corcos AF and Monaghan FV (1987) Correns, an independent discoverer of Mendelism? I. An historical/critical note.Journal of Heredity 78: 330.
Corcos AF, Monaghan FV and Weber MC (1993) Gregor Mendel's Experiments on Plant Hybrids: A Guided Study, Rutgers University Press.
Correns C (1905)Gregor Mendel's Briefe an Carl Nägeli 1866-1873. Ein Nachtrag zu den veröffentlichten Bastardierungsversuchen Mendels. Abh. Math.-Phys. Classe Kgl. Sächs. Ges. Wiss. 29:189-265.
Darwin C (1859) On the Origin of Species by Means of Natural Selection. John Murray, London, England.
Darwin CR (1862) On the two forms, or dimorphic condition, in the species of Primula, and on their remarkable sexual relations.Journal of the Proceedings of the Linnean Society of London (Botany) 6:77-96.
Darwin C (1868) The variation of animals and plants under domestication. John Murray, London.
Darwin CR (1876)The effects of cross and self fertilisation in the vegetable kingdom. John Murray, London.
Darwin CR (1877)The different forms of flowers on plants of the same species. John Murray, London.
Fisher RA (1936) Has Mendel’s work been rediscovered?Annals of Science 1: 115–137.
Hartl DL and Fairbanks DJ (2007) On the alleged falsification of Mendel’s data.Genetics 175: 975–979.
Howard JC (2009) Why didn't Darwin discover Mendel's laws?Journal of Biology 8:15.
Iltis H (1924)Gregor Johann Mendel. Leben, Werk und Wirkung. Springer, Berlin. English translation by Eden and Cedar Paul (1932), W.W. Norton & Company, Inc. New York.
Iltis A (1954).Gregor Mendel’s autobiography. Journal of Heredity 45:231-231.
Mawer S (2006) Gregor Mendel: planting the seeds of genetics. Abrams NY, Fields Museum, Chicago.
Mendel G (1866)Versuche über Pflanzen-Hybriden. Verhandlungen des naturforschenden Vereines, Abhandlungen, Brünn 4:3-47,英译本见Experiments in Plant Hybridization in Genetics: readings from Scientific American pp. 8-17. W.H. Freeman and Company, San Francisco-USA.
Mendel G (1869)Über einige aus künstlichen Befruchtung gewonnenen Hieracium-Bastarde. Verhandlungen des Naturforschenden Vereines, Abhandlungen, Brünn 8:26–31. (English translation: ‘‘On Hieracium hybrids obtained by artificial fertilisation.’’, Bateson, W., 1902 Mendel’s Principles of Heredity: A Defense. Cambridge University Press, Cambridge, UK)
Mendel G (1950) Gregor Mendel’s Letters to Carl Nägeli.Genetics 35: 1–29. 见Gregor Mendel’s letters to Carl Nägeli (1866-1873) (Translated by Leonie Kellen Piternick and George Piternick)
http://www.esp.org/foundations/genetics/classical/holdings/m/gm-let.pdf
Monaghan F and Corcos A (1986) Tschermak: a non-discoverer of Mendelism. I. An historical note.Journal of Heredity77:468-9.
Morgan TH (1909) What are “factors” in Mendelian explanations? American Breeders Association Reports 5:365-369.
Nogler GA (2006)The lesser-known Mendel: his experiments on Hieracium. Genetics 172:1-6.
Orel V (1996)Gregor Mendel the first geneticist. Oxford University Press.
Weiling F (1991) Historical study: Johan Gregor Mendel (1822-1884).American Journal of Medical Genetics 40:1-25.
阅读
Mendel G (1866)Versuche über Pflanzen-Hybriden. Verhandlungen des naturforschenden Vereines, Abhandlungen, Brünn 4:3-47,英译本Experiments in Plant Hybridization in Genetics: readings from Scientific American pp. 8-17. W.H. Freeman and Company, San Francisco-USA.
饶毅 (2008) 一意孤行的伯乐
饶毅 (2008) 达尔文的泛生论
一意孤行的伯乐
西谚云:有才方识天才,庸才仅见自己 (mediocrity knows nothing higher than itself, but talent instantly recognizes genius)。
多一些慧眼识英才的伯乐,可以使更多有特长和潜质的青年获得成长的条件,得到发挥的机会。一个好的社会希望有更多老师和资源掌握者做伯乐。
这里讲一个幕后英雄的故事,由于他有才、识才、惜才、爱才,使划时代的科学发现成为可能:
他不是科学家,却能抓住关键的科学问题;
他不在学术界,却能判断雇员的智力水平;
他不顾他人评价不同,坚决相信自己的判断;
他不顾自己资源有限,长期支持一人的研究。
没有他,很可能就没有作为科学家的孟德尔,也就不可能于1866年在学术机构以外诞生遗传学。
我在《孤独的天才》中介绍了孟德尔及其研究,可以看到:在科学界,孟德尔是孤独的。
就孟德尔的研究而言,其个人才能是必需,但非充分。因为,实验科学到一定阶段和规模,除个人的热情和才能以外,常常还需要其他条件。
那么,没有家庭背景和条件的孟德尔,是怎么成长为科学家的?如何能持续十年开展科学研究?
原来,虽然孟德尔在科学界的大环境没得到支持,但是,他在赖以生存和工作的局部环境中却获得了坚定的支持。
孟德尔做出重要发现,在研究中以其才能为主,运气成分不多。他一生最大的运气,不是科学研究过程本身,而是碰到了伯乐。
欣赏和支持孟德尔的人不止一位。但是,给予孟德尔最有力、最持久、最重要支持的,是修道院的道长纳泊(František Cyril Napp, 1792-1867)。
修道院的智力环境
纳泊于1821年到奥匈帝国的布鲁恩(Brünn,现捷克Brno)的圣汤玛斯修道院(the Abbey of St. Thomas)。这是一个天主教奥古斯丁教派的修道院。1824年,纳泊任道长,直至1867年去世。孟德尔(1822-1884)一生的遗传学研究最主要的时期(1854-1864)皆在纳泊任内。
布鲁恩离维也纳一百多公里。当时纺织工业发达,对羊毛很有需求,对水果也有较大需求。当地动物和植物育种协会较多,交流讨论频繁。协会记录显示,当地学者进行了许多育种实验,从改善品种的现实需要涉及基本科学问题。
纳泊的能干使修道院收入较好。其中,育羊是修道院重要的经济来源。但是,用实际应用的需求不能完全解释纳泊与当地动植物育种教授的密切联系。
1870年,孟德尔自己任道长时指出:“修道院从来都认为培育所有方向的科学是首要任务之一” 。这至少反映了他对前任纳泊时期修道院工作实质的认识和评价,也表明他有意愿继续此传统。
修道院不仅环境优美,而且有较好的图书馆。
纳泊吸引并支持有智力追求的神父,在人数不多(当时15位修道士)的修道院形成了一个有智力追求的群体,有革命家、作家、数学家、哲学家、语言学家、作曲家和指挥家。修道院的厨娘Luise Ondrakova后来都出版了烹饪书。纳泊主持的修道院给家庭经济状况不好的人提供了智力追求的环境。
纳泊给修道院神父们提供的优越环境,并非毫无阻力。他与布鲁恩教区主教Anton Ernst von Schaffgotsch (1804 - 1870)多次发生冲突。1854年Schaffgotsch主教受布拉格红衣主教委托巡视各修道院。他视察圣汤玛斯修道院后,非常不满纳泊过多外面的兼职、修道院其他神父过多参与科学和教育而忽略传教。他基本把Napp手下的每一个修道士都批评了一遍。对当时在维也纳大学进修的孟德尔,Schaffgotsch批评道:“他,用修道院的经费,在维也纳的世俗机构学习亵渎的科学,以期成为国立机构的同类科学的教授”。 这是Schaffgotsch对修道士最温和的批评。Schaffgotsch给上级的报告认为圣汤玛斯修道院无可挽救,建议撤销。因为纳泊已当选为终身道长,Schaffgotsch主教建议给他退休养老金,其他神父个案处理。
当时Schaffgotsch主教的报告:1855年底,布拉格红衣主教将它转交给了罗马教廷。我们不知道罗马教廷和布拉格红衣主教如何决定,也不知道纳泊如何应付,只知道圣汤玛斯修道院没有被关闭。
如果1855年关闭了修道院,那么孟德尔到哪有饭吃?他刚刚开始的实验命运如何?就都比较难说了。
纳泊爱才
人不可能十全十美,可能有性格问题,或其他问题,有才华的人也不例外。但纳泊看中人才后,看其主流,不怕其他人非议,保护他们。
在孟德尔之前,纳泊就支持过其他人。1830年,纳泊请数学家、修士Aurelius Thaler(1796-1843)在修道院建植物园,栽培稀有植物。孟德尔入修道院时,Thaler已去世三个月,但其植物园还在。当时由Franz Matouš Klácel (1808-1882)打理。
Klácel于纳泊1827年加入修道院。Klácel爱好广泛,从哲学、诗词、写作、植物到社会活动,他是修道院的两位捷克人之一(纳泊和孟德尔都是日耳曼人),鼓吹捷克独立。他曾任当地哲学教授,很受学生欢迎。1844年,Klácel因为讲黑格尔哲学而被保守的势力剥夺教授资格。纳泊帮他伸冤未果,就让他在修道院管图书馆。1848年革命活动失败后,Klácel曾鼓动包括孟德尔在内的6位神父签名要求允许他们自由教学,也没成功。1869年,Klácel移民美国,做过报纸编辑、出版商、作家,再也没做神父。
Klácel对植物感兴趣,做过植物实验,也经常和孟德尔讨论,内容包括达尔文理论发表以前的进化论雏形和后来的达尔文学说。
纳泊也曾鼓励Tomás Bratránek (1815 - 1884)继续深造,送他到w维也纳大学攻读博士学位。Bratránek 也对植物感兴趣。Bratránek 到外地做教授后,继续和孟德尔等保持通讯联系。
作曲家Pavel Křížkovský(1820-1885)是孟德尔的好友。他也来自穷人家庭,入修道院谋生。他的音乐非常受人欢迎,但Schaffgotsch主教禁止他创作和指挥非宗教音乐,只能是教会音乐。1865年,Leoš Janáček (1854-1928) 通过父亲与Křížkovský的关系,加入圣汤玛斯修道院的合唱团。圣汤玛斯修道院是Janáček的第一个机构,Pavel Křížkovský是Leoš Janáček的第一位音乐老师。Leoš Janáček以后成为捷克最重要的作曲家之一。
孟德尔做研究的关键期间,纳泊、Klácel和Křížkovský 等人形成相互支持、相互刺激的智力环境。
有深刻科学见解的神父
纳泊本人有浓厚的智力兴趣,不限于神学和哲学,也包括历史和农业。他担任过园艺协会会长、育羊协会成员。孟德尔加入的自然史学会,是农学会的分支,而纳泊曾任农学会的副会长。纳泊支持人工育种,知道杂交对实际应用的重要性。纳泊在学会刊物上发表过作物、水果和昆虫的文章。
纳泊不仅吸引一批有才华的人,提供物质保证和智力环境,而且他自己对科学有相当深刻的见解。
1836年,纳泊就曾在育种讨论会上提出:遗传就是关于动物的内在组织影响外在型式,应该研究。
1837年,纳泊在总结学会讨论时明确提出:应该讨论的问题不是培育的过程,而是遗传了什么、怎么遗传的。
这实际是遗传学的核心问题:神父纳泊,定义了尚未诞生的遗传学。
纳泊对孟德尔的超常支持
1843年孟德尔的老师将他推荐给纳泊,纳泊后来对孟德尔的支持可以说是到了一意孤行的程度。
虽然纳泊看到了孟德尔的研究结果、去世前一年见证了孟德尔论文的发表,但他并未看到学术界对孟德尔理论的认同。不仅他没看到,孟德尔本人也没看到。因此,说纳泊对孟德尔的支持是至死不渝,并非夸张。
1845年至1848年,孟德尔在神学院学习,其中也学了科学和农业技术。
1849年,27岁的孟德尔正式行教时间不长,纳泊就致信布鲁恩教区主教Schaffgotsch:孟德尔学科学非常勤奋,但很不合适做传教的神父。原因是他造访病人或有痛苦的人时,发生不可克服的羞涩。他这个问题导致他自己危险地生病。Schaffgotsch并不喜支持孟德尔,但纳泊说孟德尔适合教书,Schaffgotsch让步。修道院的募捐理论上是支持传教的,神父的首要任务是传教,不传教还可以吃修道院的饭,没有道长的庇护恐怕做不到。
孟德尔先到布鲁恩城外的Znaim教中学,为暂时不能上课的老师代课,同事学生都说他教得好。原来他的羞涩只在“传教”,不在教书。只教了几个月,老师回来了,不用他再代。纳泊又把孟德尔送到布鲁恩一所学校,又代了几个月课。
总不能永远代课。彼时奥匈帝国正式教师已需证书。孟德尔于1850年申请考教师证书,但没考过动物学和地质学部分。考官笑话他不知道动物学名,全部用德文口语。这个缺陷可能在孟德尔1866年论文中还有痕迹。他对于自己选用的豌豆,具体是何种属、学名是什么,不是非常确定。如果有现代心理医生,也许会猜他写论文时的犹豫和他以前考教师证书时说不出学名的关系。
有个考题问动物和人的关系,意思是生物学关系,孟德尔举例说:猫是能吃老鼠的有用动物,有漂亮的毛发。后来孟德尔的崇拜者看到他的答卷都觉得很可笑。幸运的是,考官客气地说:孟德尔不缺勤奋和才能,如果有机会到能接触更多信息的地方强化一下,可能会合适。
孟德尔第一次考中学教师资格考试没通过后,纳泊给主考老师之一Andreas von Baumgartner(1793-1865)写信,问为什么孟德尔没过。von Baumgartner说孟德尔自学到这个程度就不错,要是到大学进修一下更好。
于是纳泊送孟德尔到维也纳大学进修。Schaffgotsch主教批准是有条件的:孟德尔在维也纳生活得像个神父,意思是得住在修道院。但维也纳的修道院没有房间给孟德尔。纳泊仍坚持送孟德尔去,不怕他受大城市腐败的影响。纳泊也曾说:我会不惜经费使他得到进一步训练。
因此,孟德尔是因为考试不及格而获得上大学的机会。1851年到1853年,孟德尔在维也纳大学的两年学了物理、数学、植物、动物和显微镜。他修的70多个学分中,一半是物理和数学。第一学期全部学物理。第一位物理学老师是多普勒效应的发现者Christian Doppler (1803-1853),孟德尔注册了Doppler两个学期的物理,可能因为他学得较好,做过物理实验的助教。孟德尔从多普勒那里学了如何用实验解决科学问题。多普勒病逝后,第二年的物理老师是Andreas von Ettingshausen (1796-1878)也是数学家,1826年曾出版《组合数学》一书。孟德尔在维也纳大学的第四个学期修了Ettingshausen的“物理仪器应用和高等数学物理”,也就学了组合分析。这为孟德尔研究遗传提供了需要的数学,使孟德尔成为用数学成功研究生物的先驱。
维也纳大学教孟德尔的生物老师有Edward Fenzl (1808-1979)和Franz Unger(1800-1870)。Fenzl教的是保守陈旧的植物学,认为有超出物理化学原理的活力推动一切生物。Unger是开明的老师,讲新思想、新进展,他讲了达尔文以前的朴素进化思想,也介绍了Mattias Jakob Schleiden(1804-1881)的“科学植物学”,以Schleiden、Theodor Schwan(1810-1882)和Rudolf Virchow(1821-1902)等创立的“细胞学说”理解植物,认为植物整个都由细胞组成。Unger提出一个花粉细胞和一个卵细胞结合后长成植物个体,这部分内容对孟德尔做研究和分析结果有直接意义。经Unger推荐,孟德尔念过其他科学家的植物杂交实验论文。1866年,孟德尔在论文中引用的Josef Kölreuter(1733-1806)和Karl Friedrich von Gärtner (1772-1850),他都在维也纳时学过。孟德尔也学了显微镜,为他日后的植物杂交提供了实验操作训练。
从理论到实验,孟德尔都获得了良好的科学训练。虽然他后来不在教育和科研机构工作,他从事科学研究,不仅有扎实的基础,而且遵循科研规律。所以,和爱因斯坦一样,孟德尔也不是自立规矩的所谓“民间科学家”。
可是,孟德尔第二次还是没考到教师证书。与第一次考试不同,第二次考试没留下记录。一个说法是孟德尔考试怯场。另有个说法是孟德尔和Fenzl发生冲突,孟德尔回修道院做研究也是为了证明自己对,Fenzl错。这个故事很好听,但并无证据。
孟德尔的再次不第,并没有让纳泊怀疑自己的眼光,也没有动摇他支持孟德尔的决心。孟德尔终生都没能取得正式教师资格,但是,纳泊继续让孟德尔做代课老师,孟德尔到了布鲁恩实科中学(Brünn Realschule)。中学校长Josef Auspitz (1812-1889)也很支持孟德尔,让他无证教了14年的物理和自然史。Auspitz还可能很早认识到孟德尔发现遗传规律的重要性。
对以两次考试失败象征缺乏他人认可的孟德尔,纳泊的支持非但没有减少,反而变本加厉:1854年,他增加对孟德尔的支持,不仅精神支持,而且物质支持。孟德尔做实验需要暖房,纳泊就出资修建。
1848年Klácel积极参与革命活动后,将修道院的植物园交给孟德尔。为了做遗传实验,孟德尔需要暖房。这相对于一个不大的修道院来说,是很大的一笔开支。1854年,孟德尔回来不久,纳泊开始给他盖暖房,1855年交付使用。用这个暖房和一个露天的场地,孟德尔通过长年的研究奠定了遗传学基础。
虽然在科学界,没人支持、接受孟德尔,但在小小的修道院里,却有纳泊道长一如既往、尽心竭力地为他提供着强有力的支持。
纳泊自始自终全力支持孟德尔:孟德尔没钱吃饭,纳泊收他进修道院;孟德尔喜欢科学,纳泊就让他不用传教;孟德尔没有教师资格,纳泊就让他代课;孟德尔没有考过证书,纳泊让他去大学进修;孟德尔需要研究条件,纳泊就给他盖暖房。
纳泊去世前两年,孟德尔宣读了研究结果。纳泊知道孟德尔的结果和理论。他也许理解孟德尔工作的伟大,也许并不那么理解,但正是他一如既往、尽心竭力的支持成就了孟德尔,让他成为了超越时代的天才,让修道院成为了遗传学的诞生地。
纳泊是对生命科学起过最重要作用的伯乐,是世界上最有成效的伯乐之一。
http://www.mendelweb.org/
Henig RM (2000). The Monk in the Garden: The Lost and Found Genius of Gregor Mendel, the Father of Genetics. Houghton Mifflin, Boston.
Iltis H (1924).Gregor Johann Mendel. Leben, Werk und Wirkung. Springer, Berlin. English translation by Eden and Cedar Paul (1932), W.W. Norton & Company, Inc. New York.
Mawer S (2006). Gregor Mendel: planting the seeds of genetics. Abrams NY, Fields Museum, Chicago.
OrelV (1973). The scientific milieu in Brno during the era of Mendel's research. Journal of Heredity 64:314-318.
Orel V (1996).Gregor Mendel the first geneticist. Oxford University Press.
Orel V and Wood RJ (2000). Essence and origin of Mendel’s discovery.C.R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 323:1037-1041.
Orel V (2009). The “useful questions of heredity” before Mendel.Journal of Heredity 100: 421-423.
Peaslee MH and Orel V (2002). Contributions of the Members of the Augustinian Monastery in Brno, Focusing on F. M. Klácel, Philosopher and Teacher, and J. G. Mendel, Father of Genetics.20th SVU World Congress, American University, Washington, D.C. http://www.upt.pitt.edu/upt_peaslee/svu_2000.htm
Peaslee MH and Orel V (2007). The evolutionary ideas of F.M. (Ladimir) Klacel, teacher of Gregor Mendel.Biomedical Papers of the Medical Faculty of the University of Palacky Olmouc of the Czech Republic 151:151-156.
Weiling F (1991). Historical study: Johan Gregor Mendel (1822-1884).American Journal of Medical Genetics 40:1-25.
伟大科学家的重要错误:达尔文的遗传学
达尔文进化论的精髓,广为证明。不断出现的反对,从来没有撼动其根本。
但是,达尔文提出对遗传的想法,却有问题。达尔文1868年在《家养动植物变异》一书提出的“泛生假说”(Hypothesis of Pangenesis)。比较达尔文和孟德尔的想法,可以有多种讨论,特别是不同的科学思维方法。其中,有些问题并未过时,而很有现实意义。
孟德尔的遗传学研究,有高度的选择,而得出清晰的结果、推出明确的理论。而达尔文,将多个现象联系在一起,提出一个假说可以同时解释这些现象。科学研究过程中,大家遵循一个规则,所谓Occam剃刀,以最简单的理论解释实验的结果和观察到的现象。如果简单理论可以,就不用复杂理论来解释。如果用复杂理论来解释,那么复杂加复杂可以叠很多层,就很难讨论和验证。以简单理论作为基本步骤,科学虽然前进很慢,但较扎实。Occam剃刀是经验模式,并无标准公式。比如,是用简单的理论尽量解释很多的现象,还是对于所解释的现象要有所局限、有所选择?
1868年,达尔文把五类现象,代间遗传、发育、再生、植物嫁接、用进废退,全部联系在一起讨论。他提出的理论,把我们现在认识到的“细胞全能性”(全身很多细胞一直保持整套基因组)、和遗传规律混在一起。他的泛生论确实好像可以解释多个现象,但事后我们可以看到是不同性质和不同层次的现象,因为他求全,所以得出的反而是错误的理论。
达尔文提出生物体全身体细胞都产生泛子gemmules(后人亦称pangenes),进入性细胞中,这些gemmules的组合决定了性细胞内含,形成不同的性细胞,再产生不同的后代。在强调体细胞产生泛子的重要性时,达尔文说生成性原生质(generative protoplasm)要么不全在于生殖细胞,要么生殖细胞没有再生原生质,而是收集和选择泛子。
他在讨论中甚至接受了拉马克(1744-1829)的“用进废退”,而认为泛生假说能解释用进废退(比如连续多代人工地切掉一只角后牛生出来只有一只角的街头巷尾传说)。这是他的理论最可笑的部分,因为在解释遗传学的时候,达尔文放弃了自己的自然选择的进化论与拉马克主义不同的根本。达尔文说受外界影响的体细胞性状可以获得并通过gemmules进入性细胞而传代。
达尔文在获得F2代重新出现F1代不见了的隐性性状时,仅看到现象,提出所谓“回复原理”(Principle of Reversion),这并非原理,而是以新名词复述现象,把无知隐藏在名词后面。
孟德尔只研究代间遗传,不考虑其他现象。而且他仔细选择了实验对象,还选择了观察的性状。他明确说只研究子代一定相同于父本、或者母本的那些性状,而他知道有些性状,子代既不同于父本、也不同于母本,或者介于两者之间。这样,他得出的结果很干净,而他的理论很好地解释了他的结果。
1866年和1868年,两个理论发表后,他们两人在世期间,可能没人同时知道两个理论。
假设我们在当时看到孟德尔和达尔文的理论及其证据,一般并不能很简单地断定谁对。孟德尔的理论比较严谨,但他高度选择可能是优点,也恐怕导致理论不具有普遍意义。达尔文的遗传理论,解释现象较多,但怕是眉毛胡子一把抓。
在现在和未来做研究时,这样的问题,同样存在,只是一般来说,当局者迷,到以后才会恍然大悟。
在研究的早期,正确的选择范围和对象,是科学洞察力的关键之一。
达尔文当时认为体细胞的性状可以影响生殖细胞的遗传组成。后来德国生物学家魏思曼(August F. L. Weismann,1834-1914)提出germ plasma(种质)学说,种质只存在于生殖细胞中由亲代传给后代,生殖细胞可以产生体细胞,而体细胞不能产生生殖细胞,种质不受体细胞和环境影响而改变。完全摒弃了拉马克主义的基础。Ernst Mayr(1904-2005)称魏斯曼为19世纪仅次于达尔文的进化论学者。
魏斯曼做了一个简单的实验:他把小鼠的尾巴切掉,然后让他们生子鼠,他连续5代对901只老鼠进行了这一实验,没有一个后代的尾巴短了。反对魏斯曼的人会说5代不够,要更多代(而且可以无限代)的重复才能证明。但是实际上一般民间传说都是外界对一代动物影响(比如剪断猫尾巴)就遗传到下一代,所以,虽然5代实验不能代替几十代、几百代,这个结果还是完全否定了此前民间和学界不负责任的各种传说,也摧毁了获得性遗传的基础。
魏斯曼还用了人群的社会习俗作为例子:中国妇女裹脚多代并没有导致中国人小脚,而当时得代代继续裹才行;犹太儿童切割包皮没有导致犹太人天生无包皮,而得每代都环切才行。
如果从一般遗传性状上看来,以后的经验也都证明种质隔离的正确性。
但是,魏斯曼的实验很简单,而做结论时,不仅普遍化而且层次上升了。也就是说,其结论超出了其实验结果。
比如,性状不能获得性遗传,并不能否定体细胞有可能影响性细胞内的遗传物质。我们现在重新思考,可能还有问题。在基因概念一再变化的情况下,遗传不一定要用性状来看,而可以用分子来看,比如DNA、RNA、甚至蛋白质和其他分子或亚细胞器。
那么,我们是否可以重新设计实验,研究体细胞对于生殖细胞能否发生能够遗传的改变?目前热门的表观遗传学,对此有何意义?还是有其他更值得探讨的?
好像依然是问题。
魏斯曼原文见:
Weismann A (1893). The Germ-Plasm. A Theory of Heredity http://www.esp.org/books/weismann/germ-plasm/facsimile/
达尔文的相关一章抄录如下:
Darwin C (1868). The variation of animals and plants under domestication. John Murray, London.
Chapter XXVII
Hypothesis of Pangenesis
Having reflected much on some of the subjects described in the several previous chapters, I have been driven to some hypothetical conclusions, which may perhaps be worth giving. I will in the first place enumerate the leading points;
True seminal generation passes by a not much broken series, through gemmation or multiplication by buds, through fissiparous generation and the renewal by growth of large portions of mutilated individuals, into simple continual growth. The concurrence of the two sexual organs and often of two individuals is necessary in all ordinary cases of seminal generation; but the now well-known cases of 'true parthenogenesis' show that the unimpregnated ovum, passing through the sexual embryonic stages, can be developed into a perfect individual. The unimpregnated ovum also has the power of imbuing every part of the being into which it is developed with its own characters, independently of those of the male, as we see in hybrids; especially in those hybrids sprung from two species of which the one is strongly prepotent in the transmission of character over the other. So conversely it is with the male element. It is admitted by Müller and other physiologists that there is no essential difference in nature between the germs of an ovum and a bud; and Huxley (Transact. Linn. Soc. xxii, pp. 199, 210) has submitted to a rigid comparison the pseudovum (which is of the nature of a bud) of the viviparous aphis, with the true ovum of the oviparous aphis, and can discover no difference in their minute structure. It has, however, sometimes been asserted that only individuals produced from impregnated germs acquire new characters, and that these can only be perfectly transmitted by bud-propagation; but there is no such constant difference, only one in degree and frequency. In the chapter on bud-variation we have seen that not rarely individuals (phytons) produced from buds do display quite new and strongly marked characters, which can sometimes be subsequently propagated by seed; the new varieties arising from buds cannot be distinguished by any characters from seminal varieties; that such new bud varieties, though generally capable of more faithful transmission by bud-propagation than seminal varieties are by seminal propagation, yet occasionally revert, even after a long series of bud generations, to their priestine or parental character. This tendency to reversion in buds is one of the most remarkable points of agreement between bud and seminal multiplication. There is another and still more remarkable point of agreement: we have seen that a bud of one variety inserted into the stock of another variety, in some rare cases has certainly affected, as if by a kind of hybridization, adjoining buds subsequently produced from the stock. If the evidence be thought sufficient, then the dingy purple Laburnam (C. adami), and those marvellous orange trees which produce pure oranges, lemons and citrons as well as fruit of a mixed nature, have thus arisen, then undoubtedly the buds of distinct species can blend together and subsequently produce hybrids by budding, like those produced from the union of the true male and female sexual elements of distinct species.
There is, I believe, one general but hardly invariable difference between seminal generation and gemmation. Beings propagated by the former method, usually pass in the course of their development from a lower to a higher grade, as we see in the metamorphosis of insects & in the concealed metamorphosis of the higher vertabrata; but this passage from a lower to a higher stage cannot be considered as a necessary accompaniment of seminal reproduction, where we look to the kind of development of aphis amongst insects, or to that of all the higher vascular plants. In beings propagated by buds there is, as far as I know, no metamorphosis of this kind; that is, they do not pass first to a lower and then to a higher stage of development; unless indeed the scales surrounding the buds and bulbs of plants may be looked at as indicating such a passage. But beings produced from buds often advance in organization either during the act of their production or subsequently to it, as we see in the many cases of alternate generation. For I follow those naturalists who look at alternate generation as a process of budding, or, as in the case of the strobila of the medusa, as one of fissiparous multiplication. In beings produced by true generation, their metamorphoses, starting from a low grade and advancing to a higher one, no doubt stand in the closest relation, either at present or in past times, to peculiar lines of life and lead to places unoccupied in the economy of nature. Now from several previous considerations we may conclude that there is the closest agreement in nature between a germ and a bud. The concurrence of the two sexes, in the case of the germ being only an accessory, though very general, contingent. We are naturally why the germ, which before impregnation undergoes a certain amount of development ceases to progress and perishes, unless it is acted on by the male element; and why conversely the male element, which can keep alive for even four or five years in the spermatheca of female insects, likewise perishes, unless it acts on or unites with the germ, are questions which notoriously cannot be answered.
But as in the ordinary cases, the male and female elements concur in so similar a manner and degree in giving characters to the embryo, produced from their union, it is just possible that both perish unless they unite, simply from including too little protoplasm or formative (matter) for separate existence and development.
Budding is said by Műller to differ from fissiparous generation, whether spontaneous or artificial, in the new individual represented by the bud, not being at first perfectly organized, whilst the two or can hardly be said to be perfectly organized. A single cell of some of the lowest plants is capable of reproducing its kind. In the higher plants a bud or bulb with its scales has undergone development. When an animal capable of this kind of reproduction is divided into halves, the extremities are sometimes said at first to bud out; this apparently is a correct expression, for the papillae or projections at the cut extremities consist at first of undeveloped cellular matter. Hence there seems to be no fundamental distinction between gemmation and fissiparous generation; and the latter graduates from the conversion of the two halves of a being into two perfect individuals, through the renewal of a whole limb, into a mere cicatrice by which a wound is healed. We can hardly doubt that it is the same power which presides over the growth and increase of size of each part of the body during youth, and during the whole life of the animals which never cease growing: and we may believe it is the same power which presides over the continual repair of each part and tissue, as it undergoes incessant waste throughout life.
We have seen in a former chapter that some organic effects, which have been included by the older physiologists under the term ofnisus formativus together with the renewal of amputated limbs and the healing of wounds, differ in so far that new structures are thus produced, such as false membranes after inflammation or new bones in hydrocephalic skulls; but these structures though in one sense new, are formed of membrane, blood vessels, nerves and bones all resembling those proper to the species.
Turning now to Inheritance. According to the view just given, namely that even seminal generation does not differ fundamentally from the other methods of reproduction or from continual growth, we might have expected that every character belonging to every form would have been as regularly transmitted by all the methods ofreproduction as by continual growth. On this view that some organic effects, which have been included by the older physiologists under the term of nisus formativus together with the renewal of amputated limbs and the healing of wounds, differ in so far that new structures are thus produced it is not inheritance, but non-inheritance, which is the anomaly. We have seen in the chapter devoted to the subject, that a multitude of newly acquired characters, whether injurious or beneficial, whether of the lowest or highest vital importance, are often faithfully transmitted, frequently even when one parent alone is characterized.
We have seen that the very same characters will in one case be transmitted by inheritance and not in another case. Reason has been assigned for the belief that some characters are not transmitted, not from a failure of the power of inheritance, but from the conditions of life incessantly inducing fresh variability, as with grafted fruit trees and highly cultivated flowers. Other, perhaps all other, cases of non-inheritance, may be included under the principle of Reversion, by which the child tends to resembleits grandfather or more remote progenitor rather than its parents.
This principle of Reversion is the most wonderful of all the attributes of Inheritance. It frequently comes into action. What can be more wonderful than that characters, completely lost during scores or hundreds or even thousands of generations, should suddenly reappear perfectly developed, as we have seen with differently coloured pigeons and fowls when purely bred, but more especially when crossed, and as with the zebrine stripes on dunn coloured horses and in several such cases. Many monstrosities come under this same head, as when rudimentary organs are redeveloped, as in three toed horses, or when organs, which we may believe were possessed by an early progenitor, but of which no rudiment is visible in its descendants suddenly reappears as with additional mammae in a woman and a fifth stamen in some Scrophulariaceae. We have seen that reversion acts in bud-reproduction; and we occasionally see it during the growth even of individual animals, especially, but not exclusively when of crossed parentage, as in the rare cases described of fowls, pigeons, cattle andrabbits, which revert as they grow older to the colours of either parent or of some ancestor. We are led to believe, as explained in the chapter on inheritance, that every character which occasionally reappears through reversion, is present, though latent in each generation, in nearly the same way as in most or all females the secondary male characters lie latent and ready to be evolved, when the female reproductive organs are affected.
In every living creature we may feel assured that a host of lost characters lie latent and ready to be evolved under the proper conditions. How can we make intelligible and connect with other facts this wonderful and common capacity of reversion—this power of apparently calling back to life long lost characters?
We have seen in the latter chapters of this volume, that changed conditions of life, at first often causes sterility and when prolonged during several generations, induce much fluctuating variabilitythroughout the whole organization.
Sometimes changed conditions directly cause modifications in the beings propagated under them; by which I mean a considerable number of individuals are similarly modified. Again certain conditions, as intemperate living, causes an inheritable diseased condition, as with gout. In all such cases, as the germs and male elements are parts of the individuals which have been exposed to the new or injurious conditions, one may suppose that they have been in some way directly, though slightly, affected. But what shall we say about the inherited results of the use and disuse of particular organs? The domesticated duck flies less and walks more than the wild duck, and the limb bones of the tame duck have become in a corresponding manner diminished and increased in comparison with those of the wild duck: a horse is trained to certain paces, and thecolt inherits similar consensual movements: the domesticated rabbit becomes tame from close confinement; the dog intelligent from associating with man; the retriever is taught to fetch and carry, and these mental endowments are inherited. Nothing in the whole circuit of natural history is more wonderful. How can the use or disuse of a particular limb or of the brain affect a small aggregate of cells in the reproductive organs, in such a manner that the being developed from these organs inherits these newly acquired characters of either one or both parents?
I have now enumerated the chief leading points which we naturally wish to connect together by some intelligible bond. It will, I presume, be admitted that the protoplasm or formative matter, included within the germ and male element, and endowed with vitalforce, cause in seminal generation the development of each new being whose germs and buds agree, as we have seen, in structure as far as this is visible, in many remarkable attributes, as in varying inheritance, reversion, and hybridization, and lastly in their fully developed product. Hence it seems by far the simplest belief that protoplasm, identical in nature with that within the germ, collects at certain points to form buds. If this view be admitted it must certainly be extended to fissiparous generation, to the renewal of an amputated limb, to the healing of a wound and probably to continuous growth. We are thus led to believe that protoplasm of the same nature, must be diffused throughout the whole of each organic being, ready when super-abundant to form by budding new beings, both at the period of maturity and in the cases of alternate generation during youth; and ready to form new structures as after inflammation, and ready to repair lost or wasted structures. On this view we must believe that the reproductive organs do not by any means exclusively form the generative protoplasm, if indeed they form any of it, but only select and accumulate it in the proper quantity, and make it ready for separate existence.
We can thus understand the antagonism that has long been observed in plants between increase by buds, rhizomes, suckers and seminal generation (and indeed between the latter and active growth during youth); for in both cases the same protoplasmic matter is consumed; and there is not enough for both methods of propagation.
It is surprising that this antagonism should be as general as it is; but it does not invariably hold good; for the young males of the salmon, whilst very small, have their reproductive organs active; and Ernst Haeckel has recently (Monatsbericht Akad. Wiss. Berlin. Feb. 2, 1865) described the wonderful case of a medusa, with its reproductive organs active, which at the same time produces by budding a widely different form of medusa, which likewise has the power of seminal reproduction.
Furthermore, I am led to believe from analogies immediately to be given that the protoplasm or formative matter which is diffused throughout the whole organization, is generated by each different tissue and cell or aggregate of similar cells;—that as each tissue or cellbecomes developed, a superabundant atom or gemmule as it may be called of the formative matter is thrown off;—that these almost infinitely numerous and infinitely minute gemmules unite together in due proportion to form the true germ;—that they have the power of self-increase or propagation; and that they here run through the same course of development, as that which the true germ, of which they are to constitute elements, has to run through, before they can be developed into their parent tissue or cells. This may be called the hypothesis of Pangenesis.
On this hypothesis the many different parts of the structures of each individual may be compared to so many distinct organic beings, united together, but each of which propagates its own proper form. The union is far more intimate than that of flower buds or leaf-budson the same tree, or of the polypi on the same coral; but even in these cases we have some differentiation in the so-called individuals, and some parts in common; for plants have trunks and roots in common, and some kinds habitually produce two kinds of flowers; and the polypi of some corals have certain parts and the power of movement in common.
It is known that all the species in the same great class start in their course of embryonic development from the same point; and running for a time along the same path and therefore resembling each other in their earlier embryonic stages diverge in structure more and more, according as they are more and more different when grown to maturity. So we may believe it to be with the development of the gemmules thrown off from each different tissue and cell. There is hardly any greater difficulty in believing that these many gemmules may unite or cohere, each retaining its own power and qualities, into a single true germ, than in the well-known union of two species into a hybrid, and of the hybrid with another hybrid until several species are commingled in a single individual. Most of those who have closely studied hybrids and mongrels, especially M. Naudin, believe that all the characters of both parent-species are commingled, often in very unequal degrees in the unified product, but are not fused together or changed in nature like two elements in a chemical union.
We sometimes see evidence of this in the manner in which the petals of mongrel hybrid plants are finely streaked or blotched with the pure colours of the two parent-forms; and still more plainly in their reversion during successive generations, when not crossed by either parent species to the perfect character in every respect of such parents.
We must further believe that the many gemmules which together form a germ or a bud possess a marvellous mutually elective power by which they are all brought together in proper place and in due proportions. But we know that some such power resides in the male and female elements of every species; for when the germ is placed in contact with the male element of any number of closely allied species and with its own male element, the latter alone in all ordinary cases has any effect. But under certain peculiar conditions as we have seen, the male element of a distinct species is in the most marked manner elected by the germ.
We apparently see the same elective power in the law so strongly insisted on by Geoffroy St. Hilaire, de l'affinité de soi pour soi, which is displayed in so marvellous a manner in double monsters, with their corresponding and adjoining parts perfectly fused together; for if the modern view (see Carpenter and others) be correct, these double monsters originate from the early division of a single embryo, yet the complete fusion for instance of the adjoining arms of the two monsters, into one, but not of the two other arms, seems to require Geoffrey's law. But there are many other cases in which we see the same mutual attraction of homologous parts in monsters which are not double, as when the two eyes or the two legs, are united into one, with the almost perfect fusion of the corresponding bones, muscles and nerves. So again, some normal structures, which at an early embryonic age consist of two distinct organs are perfectly fused during the subsequent development into a single organ.
According to our hypothesis, this same mutual elective powerprobably comes into play in all such cases as that described by Spallanzani, who many times cut off the leg of a newt, sometimes at one joint or place and sometimes at another, and the new limb was always perfectly formed, neither too much nor too little being added. Hence we must suppose that each part or cell of the severed surface of bone, muscle and nerve, elected or attracted the different gemmules of those cells which ought to come next in due order, and these elected others, till the whole limb was exactly reproduced. To the same elective power acting under abnormal circumstances in our gemmules, we perhaps attribute the formation of each new structures false membranes after inflammation and new bones in hydrocephalic skulls.
During the indefinite multiplication of each species by seminal generation, no one believes that the protoplasm included within the germ and male elements is handed down or distributed from parents to offspring from one large primordial stock; on the contrary, it mustbe increased or multiplied during each generation. So it must be with the diffused protoplasm in those organisms which bud into an indefinite number of new individuals, and likewise in all such cases, as that, described by Bonnet, of a fresh water worm which from a mere fragment eight times successively reproduced its whole head and tail. So it must be with the separate constituent elements of the germ, that is the many gemmules or atoms of protoplasm thrown off from each tissue and cell during its development. By the multiplication and preservation in an undeveloped state of these gemmules, I account for all latent characters — those of the male latent in the female, and those which reappear in all the wonderful cases of Reversion. There is hardly any greater a priori improbability in gemmules remaining for a long time undeveloped than in fertilized seeds lying dormant for hundreds or thousands of years; nor is there much more improbability in our gemmules, in an undeveloped condition, being held down from generation to generation, being added to or growing at each generation, than is the transmission and partial development of numerous rudimentary organs; or than in the transmission of only a tendency to the production of such rudimentary organs. When a hornless breed of cattle, for instance, is greatly multiplied, it must be believed according to my hypothesis that gemmules thrown off from the cells which formed the horns in the parent-race, have been propagated through each succeeding generation, and occasionally that these gemmules from unknown causes run through the proper course of development and form horns and thus a horned beast appears by what is called Reversion. It is hardly more wonderful that these supposed latent gemmules, when undergoing development, should find their proper place and form horns, that thus the horns of a calf from a short-horned cow by a long-horned bull, should have its horns and not its hoofs affected by the male element of the bull; or that in crossing two birds with coloured and uncoloured tails, the tail of the offspring and not all the plumage [should] be affected. We have seen that it is most difficult to determine whether mutilations are ever inherited: I know of one case where an organ has been removed during several generations before its development and consequently before it can have thrown off gemmules afterwards capable of self propagation: but if mutilations are ever inherited, as has so often been stated to be the case, we could on this view in some degree understand the cause. Lastly, we can in a rough manner understand the most perplexing of all the cases of inheritance, namely how the effects of use and disuse of parts can be inherited; for the tissues and cells increased or decreased by use or disuse (but why actually existing parts should be thus affected by use and disuse I believe is not clear to physiologists) are supposed at the period of their formation to throw off gemmules endowed with all the qualities which they have acquired.
I have not as yet alluded to one most remarkable physiological fact, namely the influence of a first impregnation on the subsequent offspring of the female. The case of Lord Morton's mare has often been quoted: after producing a hybrid to a quagga, she bore two foals to a purely-bred horse. Yet these foals in their striped legs and character of hair plainly showed the quagga influence. Several analogous cases havebeen observed in crossed domestic animals. Now by our hypothesis the male element includes gemmules of every part of the parent-structures; and these gemmules have the power of self-increase and are properly diffused throughout the whole organization. We can thus see how the adult female may occasionally and under unknown favourable circumstances become, as it were, impregnated with characters from the male, and her subsequent offspring be thus affected. If the tissues of the female herself were plastic or undergoing their first development, they might, also, thus become affected. I make this remark because in the vegetable kingdom we meet with this very case: several undoubted instances of the pollen of one species or variety affecting parts of the female flower of another species or variety have been given: thus when Gallesio fertilized an orange flower with pollen from a lemon, the fruit bore stripes of lemon peel: when pale-coloured varieties of the pea and of the stock have been fertilized by pollen from darker varieties, the coats of the seeds have become coloured. Now the peel of the orange and the seed coats of the pea or stock are as much parts of the female, as the skin, hair and womb of a female quadruped. There is this difference in the two cases, that these parts in the plant have undergone a large amount of development since the act of impregnation, and have thus been affected by the differentiation and growth of certain gemmules which were contained within the pollen grains of the male.
The hypothesis, as now given, is in truth extremely complex, butso assuredly are the facts whether or no we invent some hypothesis by which they may be embraced. The gemmules thrown off from each different tissue and cell in one of the higher animals, which together form a true germ or bud, must be inconceivably numerous and minute. But reflect how minute and numerous the organic particles must be, with which the wind is tainted by certain offensive animals over miles of space, yet these particles can affect the olfactory nerves of other animals:
Not only each developed tissueas it is continually renewed is assumed by our hypothesis to throw off gemmules, but in every animal and plant there must be innumerable latent, self-propagating gemmules, ready under fitting circumstances to be developed. More than this we know that a moth during its development passes through several different moults in its caterpillar state, and through the cocoon state, and some animals pass through many more stages; and all the characters of each stage have to be transmitted and consequently at each stage gemmules have to be thrown off, preserved and multiplied, so as to be developed in the next generation into the parent cell or tissue.
In animals which undergo the process of alternate generation, at certain stages of development, all the tissues of the body break up and pass into a higher state of development, like a caterpillar when passing into the cocoon stage; and at this period the gemmules of each organ must multiply greatly so that uniting together into germ-like bodies, several individuals are produced instead of one; but each of these new individuals must include gemmules of all the earlier stages which have been passed through and of all the later stages which have to be passed through.Notwithstanding the astounding complexity of the processes implied by this hypothesis of pangenesis, yet it seems to me to comprehend the several leading facts better than any other view. On this hypothesis we may fancifully look at each animal and plant as being compounded of many beings, in the same manner as a tree or coral is compounded of many similar beings; but in neither case have these so-called beings had a separate existence. Each of these beings, or parts, is supposed to be capable of throwing off gemmules, which whilst within the organism are capable of self-increase, and which can be separately developed at the part or organ whence they were derived, and can be united, as in the case of hybrids, with other gemmules into a single germ or bud, which reproduces the complete parent form. On this view, each organic being may be looked at as a little universe, formed of a host of different self-propagating organisms, almost as numerous as the stars in heaven, and as minute as they are immense.






































