

2.5 摩尔根的学生
摩尔根不仅自己有开创性研究,也发现、支持和培养了有特色的学生。蝇室 有几个常规的学生、还有无固定座位常进常出的学生,摩尔根并不指定课题,人 人做自己的研究,自由交流,每年夏天实验室到马萨诸塞州的Woods Hole海边 度假做研究。
在遗传学研究方面,他最重要的三位学生是Alfred Sturtevant (1891-1970)、 Calvin Bridges (1889-1938)、和Hermann Joseph Muller(1890-1967)。摩尔根在哥伦比亚大学24年期间只在1909年教过“动物学导论”,学生中有Sturtevant和 Bridges。
农民的孩子Sturtevant第一篇论文是追踪自己家里马的毛色与遗传的关系。 比他大16岁、任教于哥大Barnard学院的哥哥鼓励他去图书馆查资料、也鼓励他带着问题去找摩尔根教授,摩尔根建议他将结果发表(Sturtevant,1910),并让他于1910年进自己实验室。1911年,Sturtevant突然想到基因连锁的紧密程度可以用染色体上线性排列来表示,通过重组发生频率推出两个基因之间的距离。 当天他不做大学作业,一个晚上画出了世界上第一张遗传图谱,文章在1913年发表于《实验动物学杂志》(Sturtevant,1913)。因为他的灵机一动,出了这张图谱很有用,而价格远低于近百年后几个国家出很多钱、而少动脑筋的做出的图谱。Sturtevant 也是摩尔根实验室相对比较平稳的人,他培养的学生较多,1965年总结的《遗传学简史》广为阅读 (Sturtevant,1965)。
Bridges两岁丧母三岁丧父,由祖母抚养成人。大学期间因经济原因到摩尔根实验室做技术员,从洗牛奶瓶开始,也管制造果蝇食物。但他不满足于打工,改进了研究果蝇的基本技术(包括麻醉方法、灯光等),且加入了研究的行列,他用几万张卡片,记录自己的实验结果。从眼色开始,他发现了多种突变并确定了它们的染色体定位,成为摩尔根的研究生。1913年他发现染色体不分离现象(non-disjunction) (Bridges,1913,1914),1916 年全文发表于《遗传学》杂志第一期第一页,通过研究不分离最后确证了遗传的染色体学说(Bridges,1916)。Bridges还对果蝇性决定提出了正确的理解:X染色体与常染色体的比例等于一的确定雌性、X与常染色体比例1:2的为雄性,在果蝇Y染色体其实不参与性别决定。Bridges因心脏病而英年早逝,摩尔根白发人送黑发人。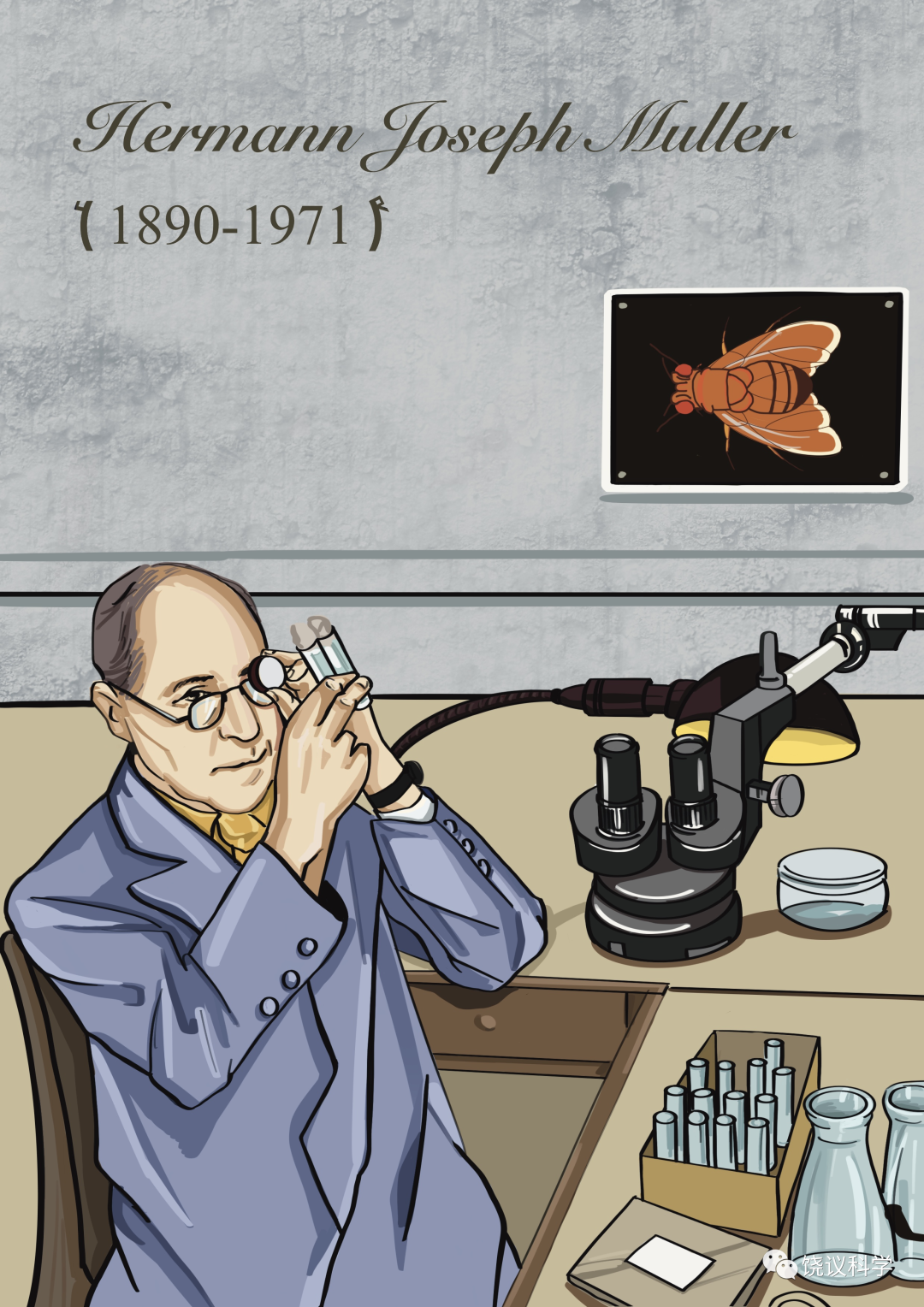
Muller通过哥伦比亚大学生的生物学俱乐部认识了Sturtevant和Bridges,得知摩尔根的研究。他在Cornell做硕士后于1912年加入摩尔根实验室完成博士论 文,独立后也用果蝇做了很多研究。
到1914年,摩尔根和学生发现的多个基因可以分成4个相连锁的组,对应于果蝇4套染色体。到1915年,他们完全可以用染色体学说解释孟德尔遗传学。 以实验和理论奠定了现代遗传学的基础后,摩尔根、Sturtevant、Muller和Bridges 四人出版了《孟德尔遗传的机理》一书(Morgan et al., 1915)。此书加上1916年Bridges的文章,使多数人接受他们的理论,虽然英国的Bateson到1916年还表示难以相信染色体是遗传的物质基础、丹麦的Johannsen到1926年才接受遗传 的染色体学说。
摩尔根1933年被授予诺贝尔奖,但当年没去领奖。引人猜想是否他后怕自己用的方法其实正是自己曾在1909年嘲笑过的孟德尔方法(数字吻合),或者因 为1933年发现果蝇唾液腺有多线染色体(polytene chromosomes)后,他的理论推导立即面临物理的检验,所以他在看到答案后于1934年才更安心地领奖。多线染色体的形成是DNA复制很多倍以后,细胞却不分裂,这样本来只有一套DNA 的单条染色体被放大很多倍而可以在显微镜下观察到。Balbiani (1881)发现昆虫唾液腺有多线染色体。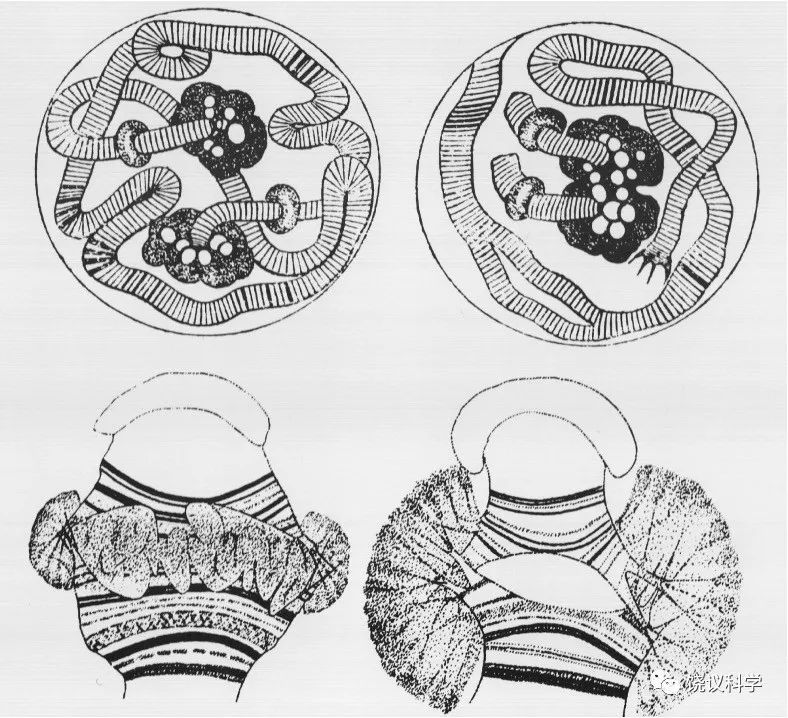
Heitz和 Bauer (1933)发现多线染色体有些区域染色深、有些浅,形成有规则的染色条带(bands)。在这样的情况下,可以猜想条带与基因的关系,能检验基因是否只是通过数字关联而推出的理论模型、还是确有物质基础。Painter (1933)发现条带密的部分与含基因多相关,而他特别重要的发现是:遗传连锁图逆转时(reversal of linkage map)染色体条带也逆转;遗传谱易位(translocation)时伴随条带易位,遗传缺失时条带也缺失。这些结果强烈地支持染色体确实是基因的物质基础。Bridges (1935、1938)更详细地描述了果蝇的多线染色体。Bridges 做的图谱,其后几十年,都是科学家们参考的标准。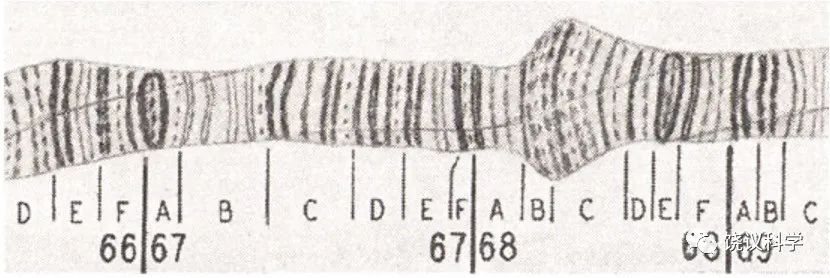
1934年,摩尔根放心地去斯德哥尔摩领奖。他将奖金分给自己、Sturtevant和Bridges三家的孩子。
2.6 摩尔根的学术传承
摩尔根通过他的科学研究、他的学生、他领导的生物系影响美国和世界的科学。
摩尔根应诺贝尔物理奖得主密立肯(Robert Millikan,1868-1953)的邀请,于 1928年创建加州理工学院的生物学部,一个小而精的系多年领先世界。他直接影响了 George Beadle (1903-1989),后者于1931至1936年在加州理工做博士后研究果蝇的遗传重组,其间(可能由摩尔根个人收入为 Beadle 提供工资)到巴黎一年研究果蝇眼睛颜色的遗传。
Beadle 研究眼色的工作起初受Sturtevant工作的影响,Beadle与Boris Ephrussi在1935和1936年从果蝇获得的结果已经开始提出基因与化学反应的关系(Beadle and Ephrussi,1936),意识到可能基因直接控制酶。
最后Beadle 在斯坦福大学与 Edward Tatum (1909-1975)进一步合作,用红色面包霉(Neurospora)研究后提出“一个基因一个酶”的概念(Beadle and Tatum,1941),开创生化遗传学,1946年Beadle 继摩尔根任加州理工生物学部主任。
摩尔根影响的著名生物学家还有1927年来自苏联的Theodosius Dobzhansky, 跟随摩尔根到加州理工学院多年,1937年出版遗传学与演化论结合的代表性作 品之一(Dobzhansky,1937)。
摩尔根实验室发现多个影响发育的基因,以后他们的学生和其他人的研究, 奠定了发育遗传学,其中最著名的是 Sturtevant 的学生 Edward Lewis 对双胸复合体的研究。未分享摩尔根奖金的Muller于1927年证明X光可以诱导基因突变、其作用与X射线的剂量相关(Muller,1927),获1946年的诺贝尔奖。他另一重要工作是 1918 年提出和制造平衡致死系,非常方便维持隐性致死突变的果蝇 品系(Muller,1918)。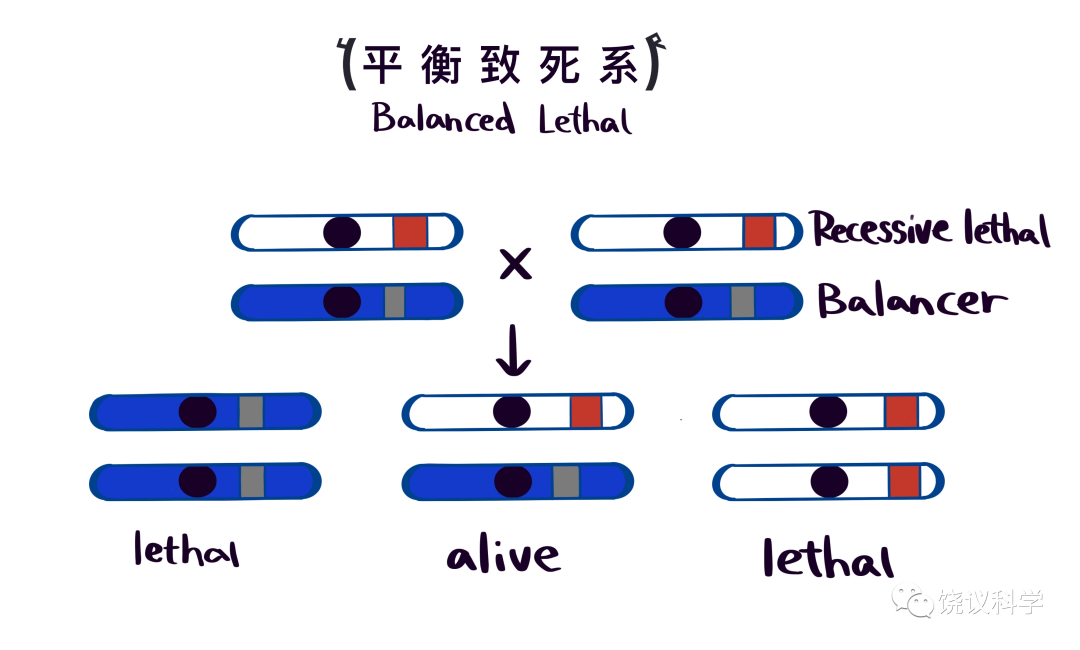
这种染色体本身不仅携带致死突变(如图中蓝色所示平衡染色体带的灰色致死突变),而且含有多重倒位(inversion),如果与另一条染色体重组也是致死,所以研究者希望跟踪的染色体与平衡染色体配对后,可以不用每代再挑果蝇,因为活着的后代都是被跟踪的染色体对平衡染色体。其他多数模式动物迄今无平衡致死系,每代需多费时间和精力,比如跟踪老鼠的隐性致死突变需要收集未交配的老鼠,割尾巴提取DNA做基因型分析,每代反复如此工作繁琐。Muller的发明近百年来省了果蝇研究者的苦力。
Muller与摩尔根关系不如其他两位学生,他比较介意功劳的归属,有个单子 写哪些是自己的想法、哪些是实验室其他人的。离开哥伦比亚大学到德克萨斯州 工作期间也与同事发生同类争论。Muller的政治立场左倾到与美国社会不融洽。 1932年到1936 年,同情共产主义的Muller在苏联科学院遗传研究所工作,李森科主义推行的伪科学流行后,Muller与之发生冲突,借口参加西班牙内战离开苏联,在西班牙期间曾与白求恩同事。到1937年,Muller既回不去苏联、也回不了美国,幸亏英国生物学家Julian Huxley的帮助转到英国。
以前邀请Muller赴苏联的遗传学研究所所长N. I. Vavilov抵制李森科主义,于1940年被捕、1941年被判死刑、1942年改为二十年徒刑、1943年死于监狱。
李森科主义泛滥的结果不仅损害了当时俄国生物学,而且俄国的遗传学和分子生物学几十年落后于 世界、今天远落后于中国。
Muller于 1940年回到美国,这时立场已是反共的Muller在美国还被怀疑亲共,从而不容易找好的教职,前几年工作不稳定,到1945年任 Indiana大学 Bloomington分校教授。他阴差阳错任教此校,影响了遗传学过渡到分子生物学。 因为欧洲不利于犹太人工作,意大利生物学家 Salvador Luria (1912-1991,1969 年诺贝尔医学奖)也任教该校,使Bloomington同时有两位重要的遗传学家。而学业不突出、起初爱好鸟类学的年轻人James D. Watson (1928-)于1947年自芝加哥大学毕业后,因为上不了加州理工学院、只能到印第安纳大学Bloomington校区念研究生, Watson说是被1946年诺贝尔奖获得者Muller所吸引。Watson到Bloomington后知道 1943年的Luria-Delbrück实验 (Luria and Delbrück,1943),学了细菌遗传学,成为Luria的第一位研究生,加入了当时以Delbrück和Luria为核心的新兴的噬菌体小组(The Phage Group),毕业后Watson先北欧后英国,1953年与Francis Crick 提出DNA的双螺旋模型,他的遗传学背景来自印第安纳大学。
摩尔根的妻子也是科学家。Lilian V Morgan (1870-1952)的家谱可以追踪到“五 月花号”,她于1887年入Bryn Mawr女子学院,1891年毕业,EB Wilson介绍她 认识摩尔根。她到欧洲留学一年后回Bryn Mawr,从1894年至1906年,她独立发表多篇发育和再生方面的研究论文。1904 年三十四岁时,她与三十八岁的摩尔根结婚,婚后继续研究再生。从1906年第一个孩子出生后到第四个孩子幼小 时期,她虽有佣人,但她在家。1921年第四个孩子十岁后,五十岁的她重返研究一线,在摩尔根实验室独立做果蝇研究。从1906年到1922年间隔16年无论文后,她从1922年至1947年再发表十二篇论文(Keenan,1983;Sturtevant,1965)。其中1921年2月12日发现一种突变的雌果蝇,经过分析,确认其两个X染色体相连 (attached X),摩尔根夫人1922年发表文章报道此果蝇,成为果蝇遗传学一个工具。
摩尔根学派对中国有直接影响。摩尔根在女子学院的研究生Alice Boring (博爱礼)于1923年至1950年任教燕京大学,不仅影响中国的生物学,且影响协和医学院通过燕京生物系入学的医学生。1922年陈祯在摩尔根实验室学习,1926 年李汝祺在 Bridges 指导后获博士学位、陈子英在 Sturtevant 指导后获博士学位。李汝祺 (1927)研究了Notch 基因突变,确定其致死期为胚胎。李汝祺回国后将自己在燕京大学的硕士生谈家桢推荐到摩尔根实验室,谈于1936 年在 Dobzhansky指导后获博士学位。李汝祺长期任教于燕京大学和北京大学,为中国遗传学的奠基人。
Muller在五十年代初期伸手救援过受李森科主义迫害的李景钧,李从北大农 学院农学系合并到中国农大后,遭当时的党总支书记迫害,出逃香港后无身份证明,Muller个人提供信件并赴香港带李见美国领事馆人员,使李能赴美任教。
当然,今天在中国工作的遗传学家与摩尔根学术谱系有关的科研人员不仅限于北大,而有更多。
节选自:饶毅 《生物学概念与途径》