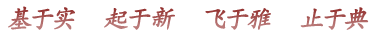
15 分子生物学筛选
15.1 按表达时间寻找基因
15.2 按表达空间找基因
15.3按功能特点寻找基因:蛙卵表达克隆
15.4 按功能特点寻找基因:钙信号检测
科学研究可以是聚焦一个科学问题、自然现象;也可以是科学家偏好一种技术,利用技术寻找可以由技术帮助解决的问题。
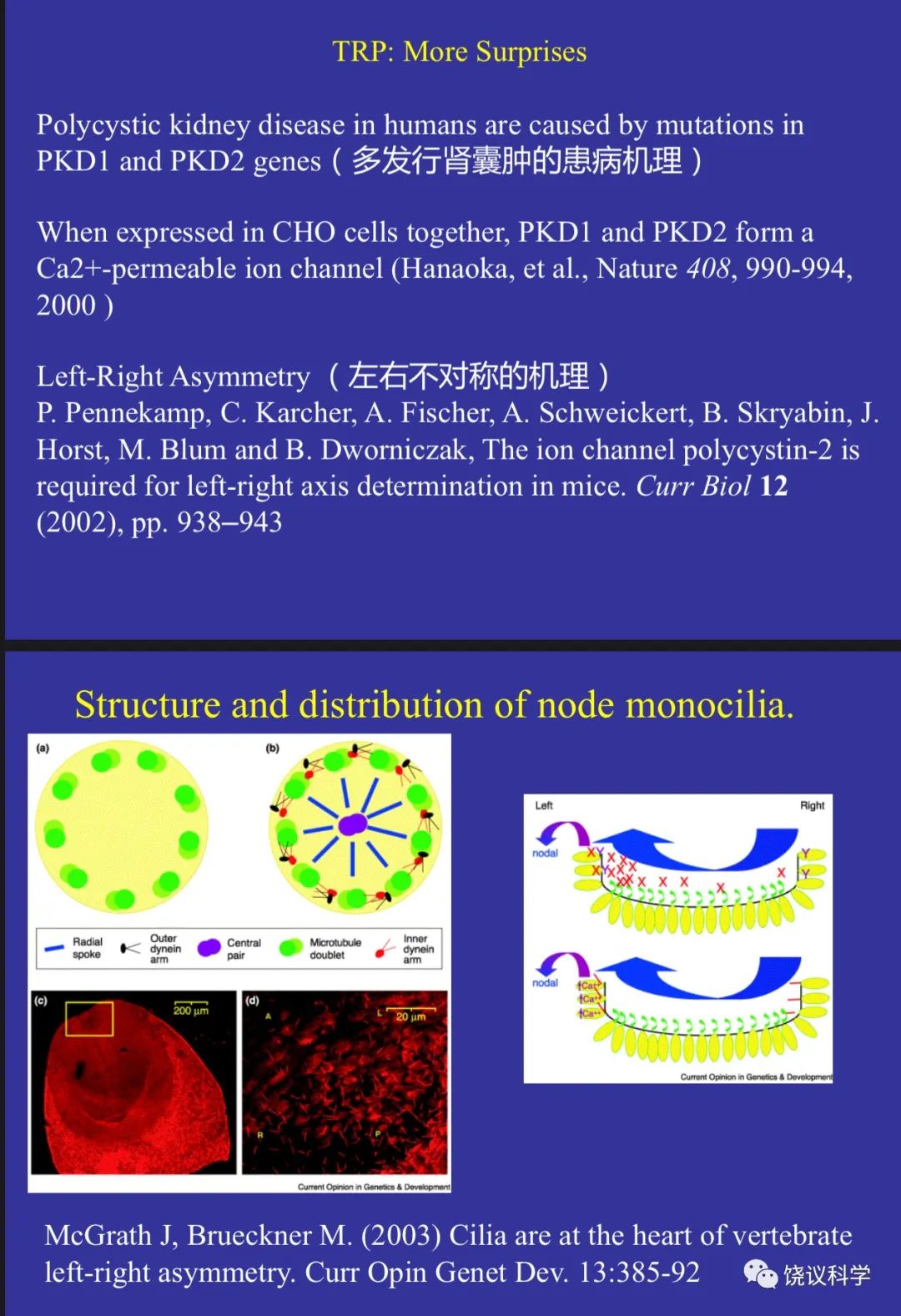
钱永健(Roger Tsien,1952-2016)以其化学和生物学背景,不断发明技术,用于生物学研究,带动技术发明逐渐成为生命科学的研究生涯。他发明钙离子检测的方法有特别的贡献。他先通过改造钙离子的螯合剂,使之在结合钙离子后发出荧光,从而用荧光强度变化检测活细胞内的钙离子浓度,有fura-2等一系列检测钙离子浓度的钙染料(Tsien, 1980, 1981)。在下村脩(Osamu Shimomura,1928-2018)(Shimomura, Johnson and Saiga, 1962; Morise, Shimomura, Johnson and Winant, 1974)和Douglas Prasher(1951-)(Prasher et al., 1992)工作的基础上,钱永健参与证明单个蛋白质(绿色荧光蛋白, GFP)可以因为结合钙离子后发出荧光(Heim, Prasher and Tsien, 1994;Chalfie et al., 1994)。
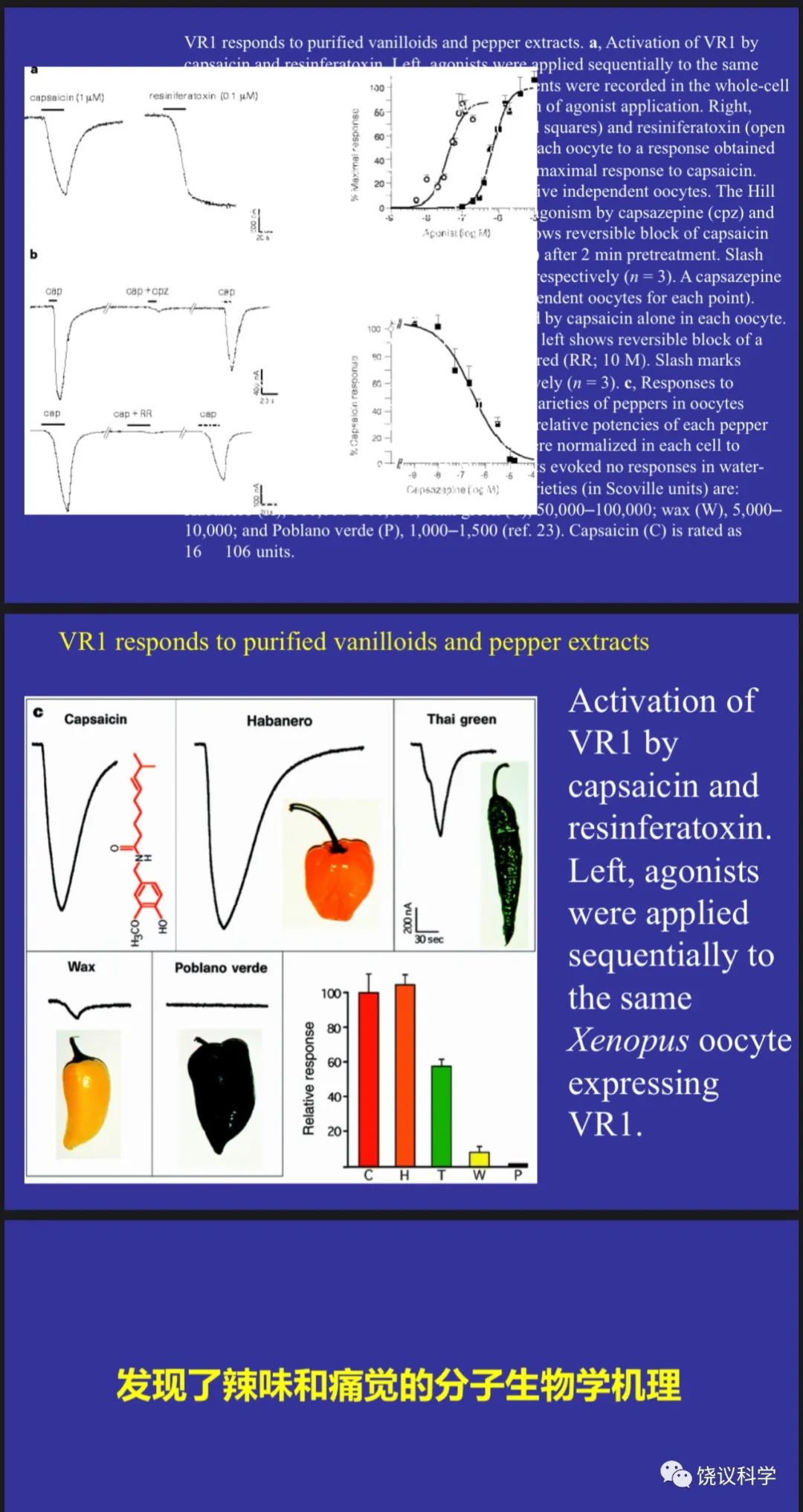
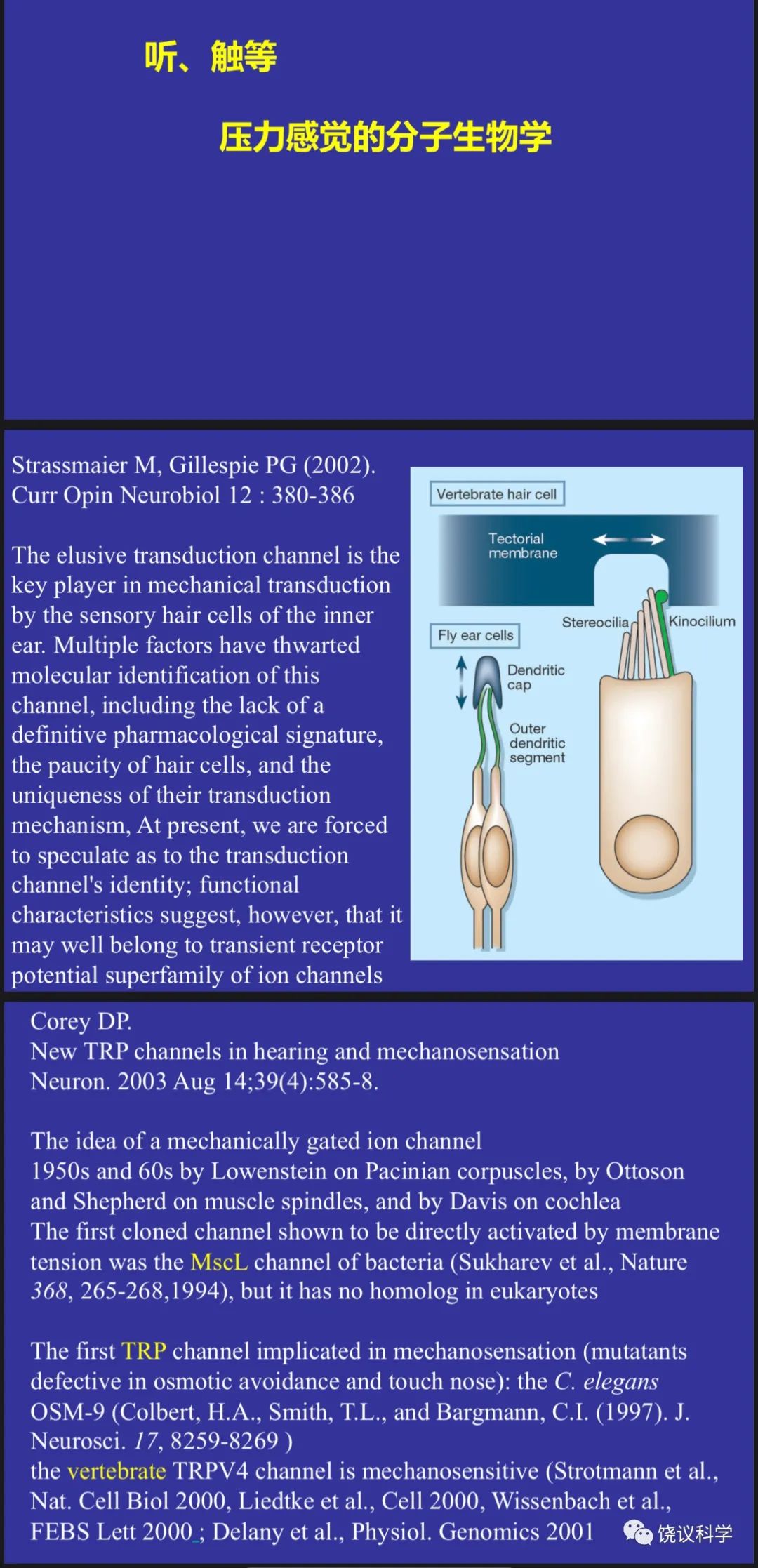
已知辣椒里面的分子辣椒素(capsaicin)可以增强痛敏感性、也可以造成热感。David Julius(1955-)于1980年代在Richard Axel实验室做博士后的阶段用表达克隆寻找五羟色胺的受体(Julius et al., 1988)。到旧金山加州大学任教多年后,他继续用表达克隆,其中一个课题为寻找辣椒素的受体,认为找到辣椒素的受体可能有助于理解痛觉的机理。这一研究最后对感觉系统的分子机理有很大影响,且不限于外周感觉系统,虽然对理解痛觉影响相对较小。已知脊髓背根神经节(dorsal root ganglia,DRG)不仅传入多种外周感觉,而且对辣椒素有反应。英国科学家用同位素标记的离子证明辣椒素(capsaicin)可以诱导大鼠背根神经节(dorsal root ganglia, DRG)的神经元产生钙离子内流和钠离子内流、不诱导氯离子外流(Wood et al., 1988)。依据这一特征, Julius带领实验室,用DRG的mRNA制备cDNA文库,然后转染到普通培养细胞(常用的HEK293细胞)。起步估计用了含一万六千多种cDNA的库,不断分群并检测,最后拿到一个cDNA,单独转染到HEK细胞就可以对辣椒素出现钙离子内流的反应(Caterina et al., 1997)。编码的蛋白质当时命名为VR1,它不仅介导对辣椒素的反应,而且介导对热的反应,被加温所激活。
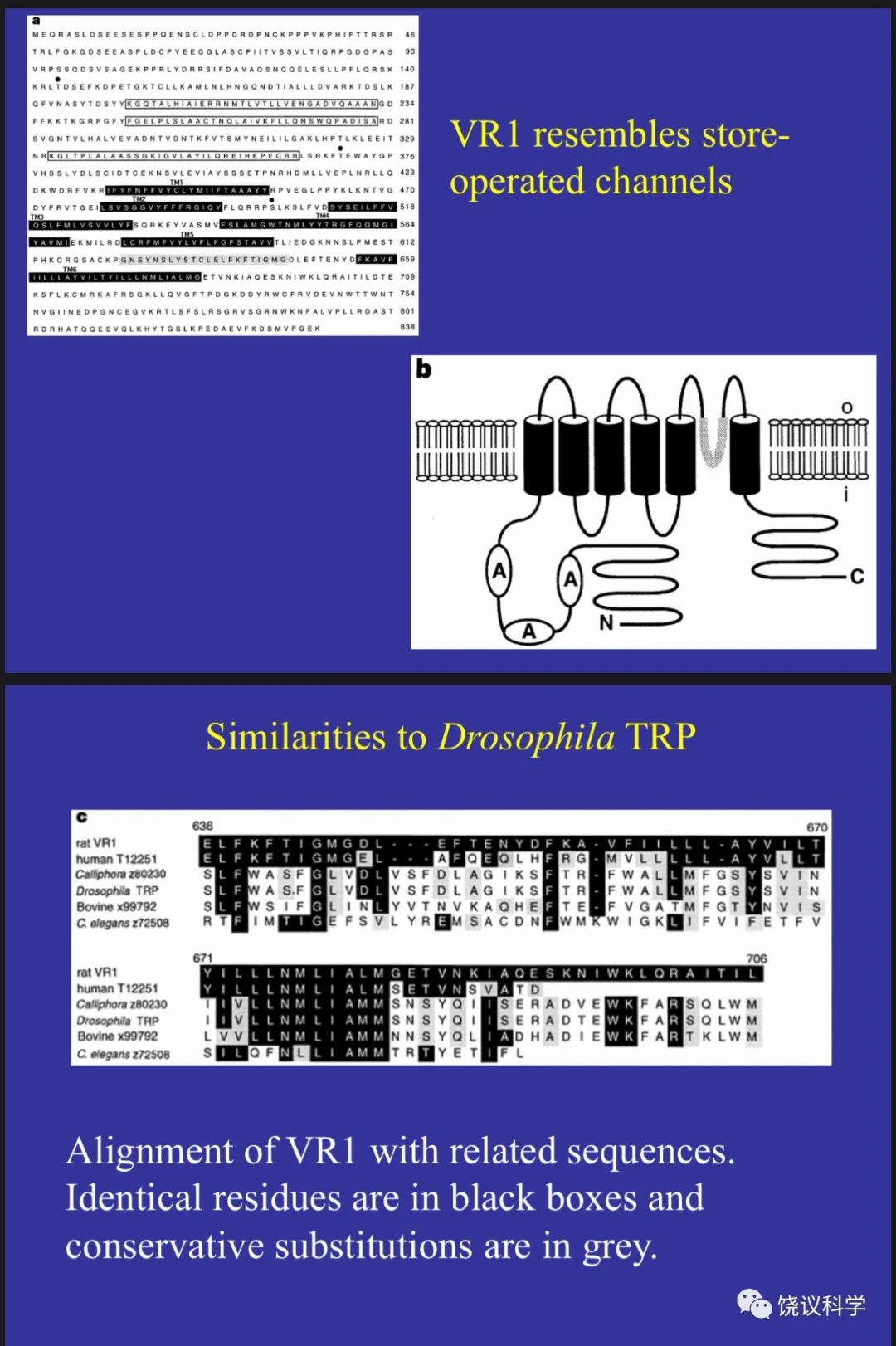
有趣的是,VR1并非独一无二的蛋白质,而是属于一个已经知道蛋白质氨基酸序列的家族:Trp通道。但是,在Julius发现VR1介导对辣椒素和热反应时,Trp通道家族的功能其实不很清楚。1969年,美国印地安纳州的Purdue大学生物系韩裔教授Willam Pak(1932-2023)用果蝇的视网膜电图(electroretinogram,ERG)研究果蝇的视觉、并寻找影响果蝇视觉的突变种(Pak et al., 1969)。同年,英国爱丁堡大学的科学家发现一种突变,根据ERG的变化,他们称突变果蝇为“瞬间感受器电位”(Transient Receptor Potential,Trp),相应突变的基因也就是果蝇的Trp基因(Cosens and Manning, 1969)。1989年,美国加州大学伯克利分校Gerald Rubin(1950-)实验室的博士后Craig Montell(1955-)克隆了果蝇的Trp基因(Montell and Rubin,1989)。他们发现Trp基因编码的蛋白质有多重跨膜区,各由大约19个氨基酸组成的疏水段。因为它的突变影响果蝇视网膜对光反应,所以最容易推测的功能是离子通道。但证明它是离子通道花了相当长的时间。Trp类似的基因后来发现于多种生物,包括人。但不仅果蝇的Trp本身并不感受光,其它的Trp具体起什么作用,如何起作用,也都不清楚。
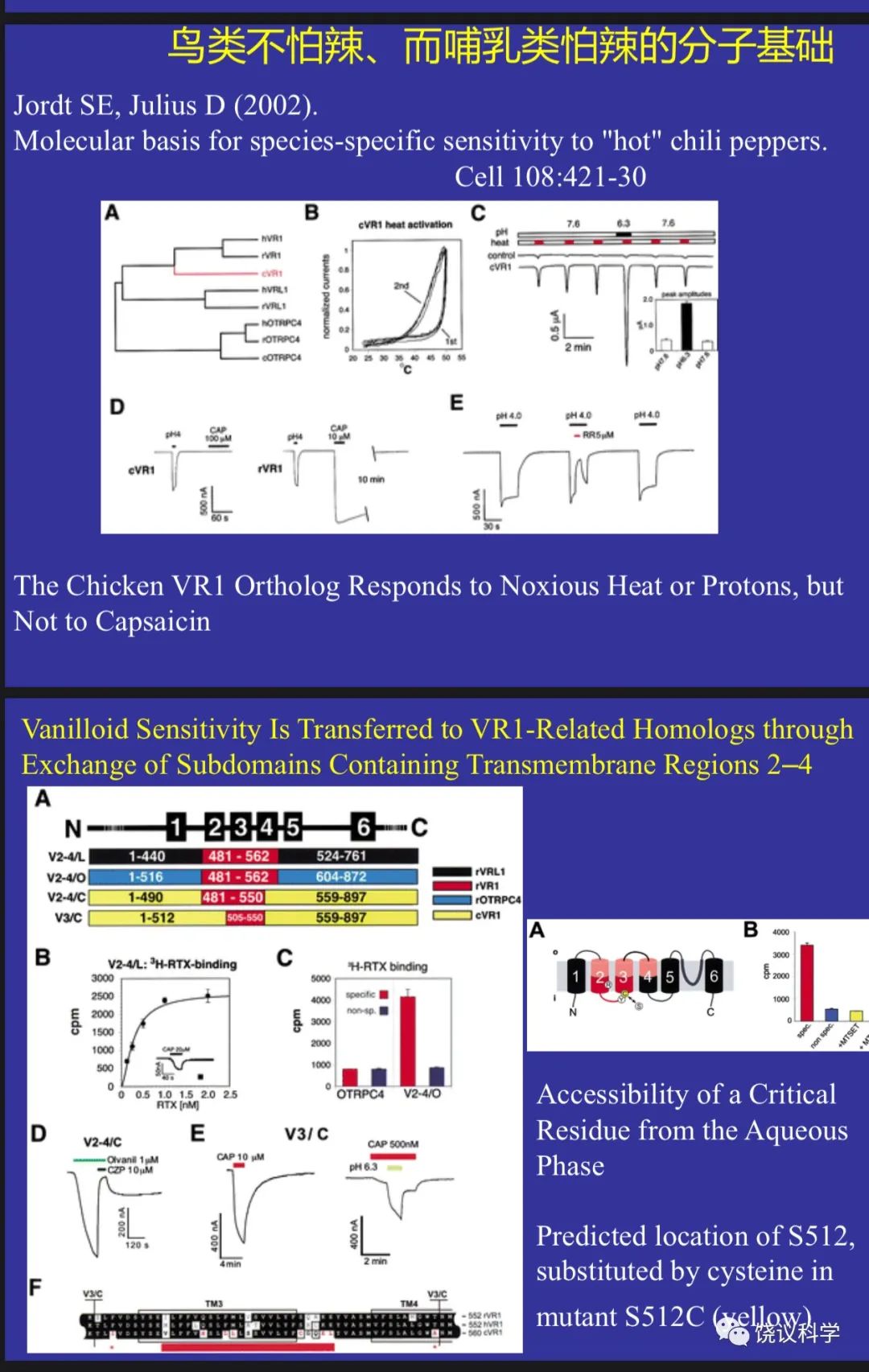
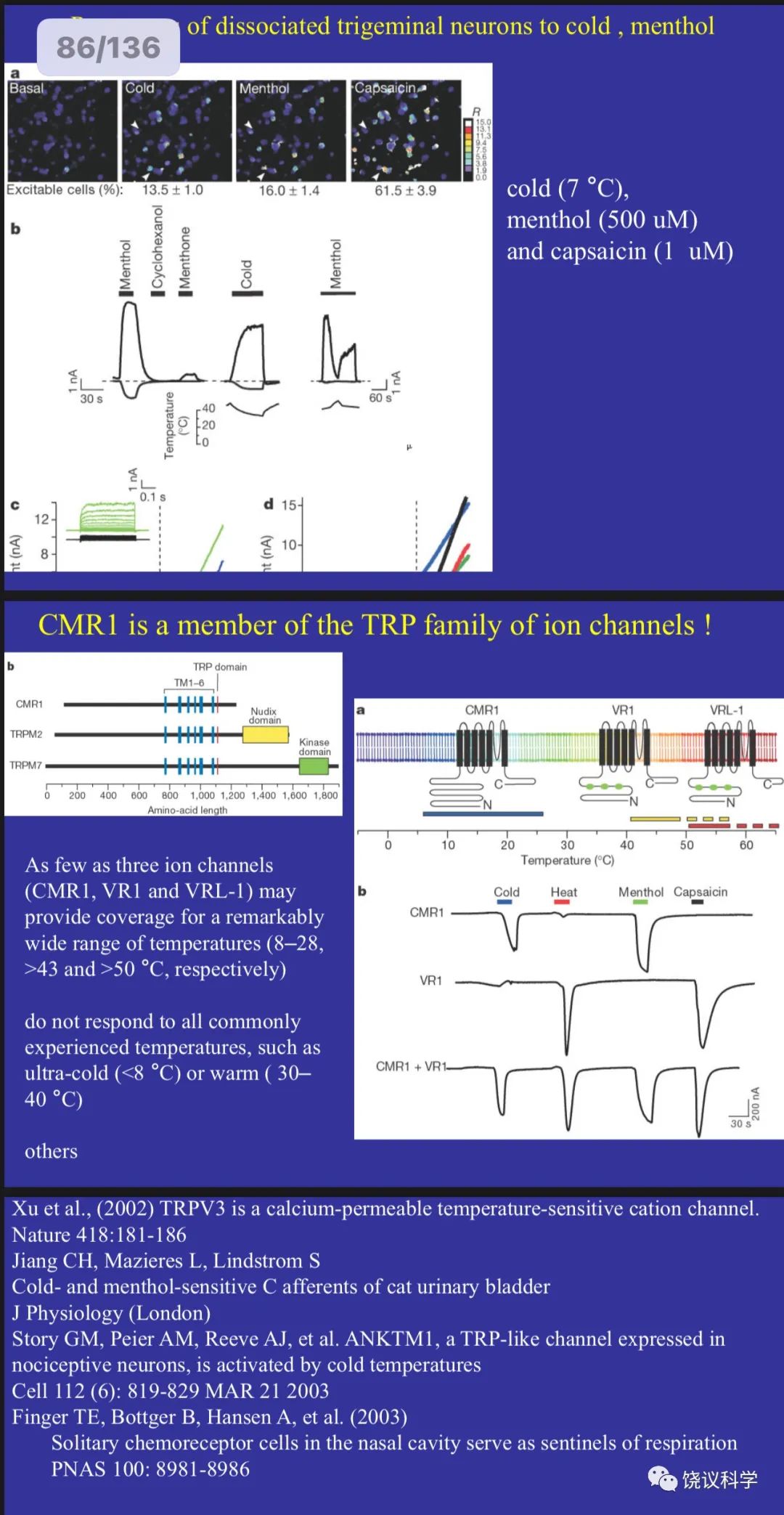
Julius实验室1997年的发现揭示了其直接功能,由此Trp的研究热火朝天,很多人加入Trp研究的行列,发现更多Trp家族成员,发现更多功能,如Julius实验室2002年用薄荷寻找冷敏感的分子(McKemy et al., 2002),结果发现是Trp家族另一成员,有多个Trp通道,温度变化的方向和范围不同的激活不同的Trp通道。还有科学家发现Trp通道感受压力,等等。
视觉、嗅觉、味觉的第一级感受分子都是GPCR。视觉的第一级是通过生物化学分离纯化最早在19世纪得到视紫红质。嗅觉的第一级也被发现是G蛋白偶联的通路,在发现对嗅觉分子敏感的腺苷酸环化酶(Pace et al., 1985)、嗅觉神经元特异的G蛋白(Jones and Reed,1989)和环核苷酸激活的离子通道(Dhallan et al., 1990)的基础上,美国哥伦比亚大学的Richard Axel(1946-)和他的博士后Linda Buck(1947-)通过多聚酶链反应(PCR)发现了小鼠嗅上皮的数以百计的GPCRs(Buck and Axel,1991),以后证明它们是感受嗅觉的分子。味觉再模仿嗅觉也得到类似发现,但味觉的感受分子只有几个,分别对应不同的味觉(甜、咸、苦、酸、鲜)(Hoon et al., 1999; Chandrashekar et al., 2009)。
确定Trp通道的功能,解决了其他常见感觉(温、部分的压、部分的痛)第一级的分子机理,有较大意义。
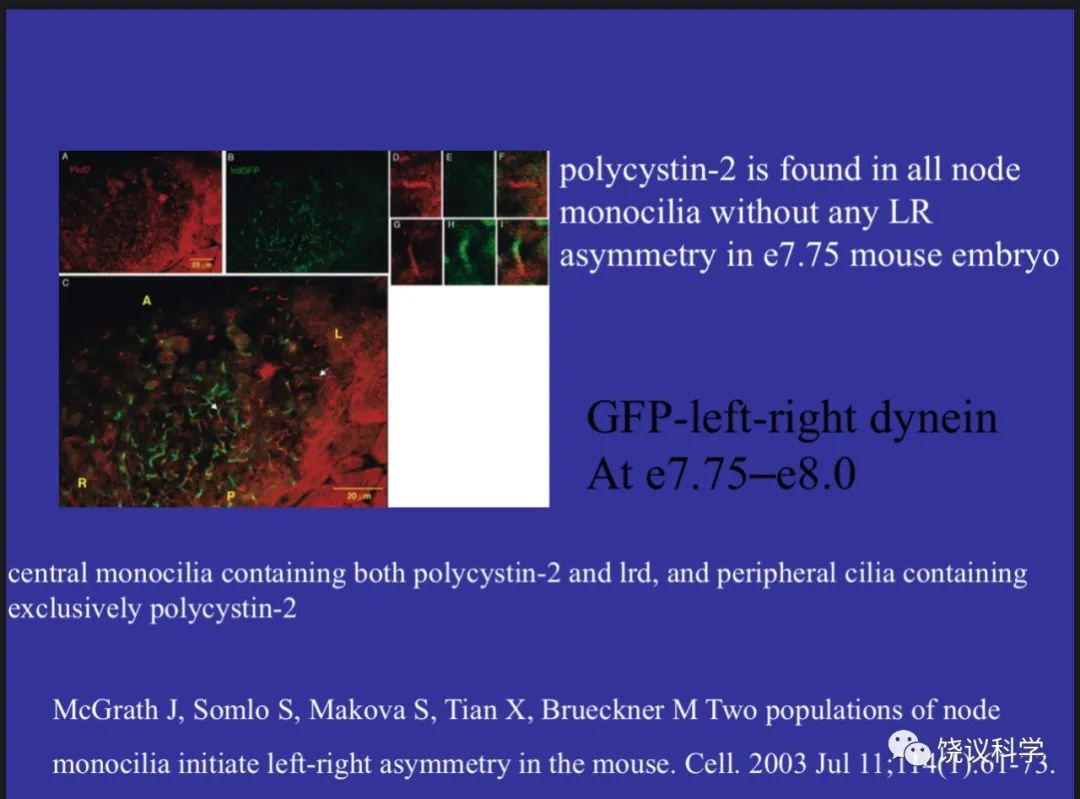
最初以为听觉也是用一种Trp通道,后来发现听觉的毛细胞还有其他结构完全不同的压力感受分子,如TMC(Kurima et al., 2002; Vreugde et al., 2002; Jia et al., 2020)。而外周和内脏的压力感受器也有多种,如美国加州的Scripps研究所的Patapoutian实验室发现的Piezo(Coste et al., 2010, 2012)和GPCR(Xu et al., 2018)。看起来简单的压力感受,却用了很多结构不同的分子。
Buck L and Axel R (1991). A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell 65:175-187.
Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD and Julius D (1997). The capsaicin receptor: a heat-activated ion channel in the pain pathway. Nature 389:816-24.
Chandrashekar JC, Yarmolinsky D, von Buchholtz L, Oka Y, Sly W, Ryba NJP and Zuker CS (2009). The taste of carbonation. Science 326:443-445.
Cosens DJ and Manning A (1969). Abnormal electroretinogram from aDrosophila mutant. Nature 224: 285-287.
Coste B,Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, Dubin AE and Patapoutin A (2010). Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 330:55–60.
Coste B, Xiao B, Santos JS, Syeda R, Grandl J, Spencer KS, Kim SE, Schmidt M, Mathur J, Dubin AE, Nontal M and Patapoutian A (2012). Piezo proteins are pore-forming subunits of mechanically activated channels. Nature 483:176-181.
Dhallan RS, Yao KW, Schrader KA and Reed RR (1990). Primary structure and functional expression of a cyclic nucleotide-activated channel from olfactory neurons. Nature 347:184-187.
Hoon MA, Adler E, Lindemeier J, Battey JF, Ryba NJP and Zuker CS(1999). Putative mammalian taste receptors a class of taste-specific GPCRs with distinct topographic selectivity. Cell 96:541-551.
Jia Y, Zhao Y, Kusakizako T, Wang Y, Pan C, Zhang Y, Nureki O, Hattori M and Yan Z (2020). TMC1 and TMC2 proteins are pore-forming subunits of mechanosensitive ion channels. Neuron 105:310-321.
Jones DT and Reed RR (1989). Golf: an olfactory neuron specific-G protein involved in odorant signal transduction. Science 244:790-795.
Jordt SE and Julius D (2002). Molecular basis for species-specific sensitivity to “hot” chili peppers. Cell 108:421-30.
Kurima K, Peters LM, Yang Y, Riazuddin S, Ahmed ZM, Naz S, Arnaud D, Drury S, Mo J, Makishima T et al. (2002). Dominant and recessive deafness caused by mutations of a novel gene, TMC1, required for cochlear hair-cell function. Nature Genetics 30:277-284.
McKemy DD, Neuhausser WM and Julius D (2002). Identification of a cold receptor reveals a general role for TRP channels in thermosensation. Nature 416:52-8.
Montell C and Rubin GM (1989). Molecular characterization of theDrosophila trp locus: a putative integral membrane protein required for phototransduction. Neuron 2:1313-1323.
Pace U, Hanski E, Salomon Y and Lancet D (1985). Odorant-sensitive adenylate cyclase may mediate olfactory reception. Nature 316:255-258.
Pak WL, Grossfield J and White NV (1969). Nonphototactic mutants in a study of vision of Drosophila. Nature 222:351-354.
Vreugde S, Erven A, Kros CJ, Marcotti W, Fuchs H, Kurima K, Wilcox ER, Friedman TB, Griffith AJ, Balling R et al. (2002). Beethoven, a mouse model for dominant, progressive hearing loss DFNA36. Nature Genetics 30:257-258.
Wood JN, Winter J, James IF, Rang HP, Yeats J and Bevan S (1988). Capsaicin-induced ion fluxes in dorsal root ganglion cells in culture. Journal of Neuroscience 8:3208-3220.
Xu J, Mathur J, Vessières E, Hammack S, Nonomura K, Favre J, Grimaud L, Petrus M, Francisco A, Li J, Lee V, Xiang F-L, Mainquist JK, Cahalan SM, Orth AP, Walker JR, Ma S, Lukacs V, Bordone L, Bandell M, Laffitte B, Xu Y, Chien S, Henrion D and Patapoutian A (2018). GPR68 senses flo and is essential for vascular physiology. Cell 173:762-775.
15.5 按功能特点寻找基因:重组激活基因的发现
第七章我们讲到,体细胞DNA重组是免疫球蛋白多样性的最重要原因。
那么,发现重组酶就应该是理解其机理的关键。
1984年,美国分子生物学家David Baltimore(1938-)当时在MIT实验室的研究生Susanna Lewis设计了实验,用k轻链基因的部分序列重构V-J重组。他们选取了鼠胚Vk和Jk及其附近的DNA序列,装在病毒里面。将这种病毒感染淋巴细胞样的体外培养细胞系PD。他们在序列Vk和Jk之间,装了来自细菌的可以产生对霉酚酸起抵抗性的黄嘌呤-鸟嘌呤磷酸核糖转移酶(xanthine-guanine phosphoribosyl transferase,gpt)。因为预计重组导致其间DNA序列翻转,所以放到病毒的是反向的gpt序列。一旦发生Vk和Jk之间重组,其间的序列翻转,gpt成为正向而可以表达GPT蛋白质,细胞能够抵抗霉酚酸。Lewis等成功地观察到,她们的分子构造,转染入PD细胞系后,经过霉酚酸后得到的细胞株多数都是因出现了Vk和Jk之间DNA重组、gpt序列翻转(Lewis et al.,1984)。
Baltimore实验室的另一位研究生David Schatz用类似Lewis的方法,再构建了一个含B潮霉素抗性基因(hph)的病毒。用两种病毒,他们检验了多种B细胞前体细胞样的细胞系,多数能够重组Vk和Jk之间DNA、翻转gpt或hph抗性基因序列,导致细胞对霉酚酸或B潮霉素有抵抗性。但如果用同样的病毒感染3T3成纤维细胞系,就不能发生Vk和Jk之间DNA重组(Schatz and Baltimore,1988)。Schatz在实验室周会上讲自己工作进展时,以色列来的博士后Yoav Citri(1953-1995)建议想办法激活3T3细胞中的V(D)J重组活性,这一建议影响了后面的研究(Schatz and Baltimore,2004)。
要在3T3细胞发生Vk和Jk之间DNA重组,当时一般的设计应该是把可重组Vk-Jk的细胞(前B细胞)的mRNA做成cDNA文库。文库的大量不同cDNA转染到3T3细胞,后者含有可以检测Vk和Jk之间DNA重组的gpt或hph。在没有转染来自前B细胞cDNA的情况下,3T3细胞会被霉酚酸或B潮霉素杀死。而如果有前B细胞的cDNA导致Vk和Jk之间DNA重组,使得其间的gpt或hph基因成为可以表达的方向,从而有GPT或HPH合成,这样的3T3细胞就能抵抗霉酚酸或B潮霉素。多数3T3细胞都被转染了与重组酶无关的cDNA,这些3T3细胞会被霉酚酸或B潮霉素杀死,剩下可抵抗霉酚酸或B潮霉素的3T3细胞就很可能是表达了重组酶的cDNA从而导致Vk和Jk之间DNA的重组。
因为一种细胞表达很多mRNA从而可以得到很多不同的cDNA,如果是一种cDNA编码重组酶,那么这种方法一般是可行的。但是,如果是多种cDNA编码的蛋白质才能引起Vk和Jk之间DNA重组,那么同时含有两种或多种起作用的cDNA的几率就会很小。Baltimore当时在Whitehead研究所的同事Robert Weinberg(1942-)用分子生物学研究癌症,Weinberg实验室用基因组DNA制备文库。将这种文库转染到3T3细胞时,每个3T3被转染的不会只是单个cDNA,而是一段基因组DNA,可能含相邻几个基因。
Schatz把B淋巴瘤的基因组DNA转染到3T3,基因组DNA接了一种抗性基因(组氨醇脱氢酶),首先用这种抗性基因针对的药物(组氨醇)选择被B淋巴瘤的基因组DNA转染了的3T3细胞。然后用霉酚酸选择发生了V-J重组的3T3细胞。他们从1500个抵抗组氨醇的3T3细胞克隆得到1个抗霉酚酸的3T3细胞株(Schatz and Baltimore,1988)。他们分析发现这一3T3细胞株确实出现了Vk和Jk之间DNA重组,使得其间的gpt表达,而不是其他原因对霉酚酸出现抵抗。但很多人不信,他们附近的实验室认为他们的结果不可靠、发表的是伪迹(Schatz and Baltimore,2004)。
Schatz与研究生Marjorie Oettinger组成两人合作,每天倒班、想方设法克隆重组激活基因(RAG)。他们用特定的寡核苷酸标记基因组的DNA,从出现重组的细胞中通过用于标记的寡核苷酸取得基因组DNA片段,以此克隆了RAG的催化亚基RAG-1(Schatz, Oettinger and Baltimore,1989)。RAG-1基因编码含一千余氨基酸残基的蛋白质,其同源基因存在于有V(D)J重组的动物如人、鼠、兔、羊、马、鸡、蛙。在人和鼠能够进行V(D)J重组的细胞系,RAG-1也有表达。RAG-1表达的细胞和组织与V(D)J重组活性相关:RAG-1表达于鼠的胸腺、骨髓和胚胎,而不在脾脏和脑表达。
在一派大好的形势下,有个异数:含RAG-1基因的18kb DNA,催化V(D)J重组的活性居然与最初全基因组切的片段效率一样,比预计的低100到1000倍。当时他们的估计是也许因为所用的基因组DNA不够长,缺乏某些未知的转录控制元件和34个氨基酸(Schatz, Oettinger and Baltimore,1989)。但后来发现,得到全长的cDNA后,活性也不高(Oettinger et al.,1990)。他们经过几个月的迷惑不解,不断提各种可能。其中一个比较难以置信的可能性是RAG-1旁边碰巧还有一个基因,与它共同起作用(Schatz and Baltimore,2004)。在细菌中,已知相关功能的基因聚集在附近,而动物细胞的基因早已知道并非如此。
她们发现在原初有重组活性的18kb基因组DNA中,RAG-1基因只占其中6.6kb,另有11kb,余下的基因组DNA中还有一个基因。她们进一步的工作证明这个基因可以提高RAG-1重组效率1000倍,因此她们称这一基因为RAG-2(Oettinger et al.,1990)。以后知道,RAG-1独立进化自转座酶Transib,后来又捕获了RAG-2基因进一步增强其活性,之后RAG-1和RAG-2的序列和功能变化成为专门用于Ig基因重组的酶(Carmona and Schatz,2017)。
Carmona LM and Schatz DG (2017) New insights into the evolutionary origins of the recombination-activating gene proteins and V(D)J recombination. Federation of European Biochemical Societies Journal 284:1590-1605.
Lewis S, Gifford A and Baltimore D (1984) Joining of Vk to Jk gene segments in a retroviral vector introduced into lymphoid cells. Nature 308:425-428.
Oettinger MA, Schatz DG, Gorka C and Baltimore D (1990) RAG-1 and RAG-2, adjacent genes that synergistically activate V(D)J recombination. Science 248:1517-1523.
Schatz DG and Baltimore D (1988) Stable expression of immunoglobulin gene V(D)J recombinase activity by gene transfer into 3T3 fibroblasts. Cell 53:107-115.
Schatz DG, Oettinger MA and Baltimore D (1989) The V(D)J recombination activating gene, RAG1. Cell 59:1035-1048.
Schatz DG and Baltimore D (2004) Uncovering the V(D)J recombinease. Cell S116:S103-S106.
附:2000至2003年讲课ppt
2013年:http://blog.sciencenet.cn/blog-2237-749393.html