

Nat Genet | 张浩岳/Gerd Blobel合作揭示凝缩蛋白缺失的有丝分裂染色体中基因组折叠的机制
哺乳动物基因组具有高度的三维组织结构,包括形成A/B区室、拓扑关联域(TAD)和染色质环【1】。在细胞周期中,所有结构特征都是高度动态的【2-5】。进入有丝分裂后,A/B 区室、TAD 和染色质环被破坏,同时伴随凝缩蛋白复合物(condensin)的加载【6】。尽管有这些显著的变化,但在有丝分裂染色体上仍然可以检测到弱结构域样的结构【2】。这些残留的有丝分裂折叠模式背后的机制尚不清楚。
在凝缩蛋白缺失的细胞中,A/B 区室和 TAD 的消失在进入有丝分裂过程中延迟【6,7】。在形态学上,凝缩蛋白的消耗消除了有丝分裂染色体的圆柱形构型,并触发了单个染色单体的混合,但有丝分裂染色体的总体积基本保持不变【7】。这些和其他观察的结果表明,除了凝缩蛋白之外,其他力量也可能影响有丝分裂染色体的构象【6】。因此,研究凝缩蛋白缺陷有丝分裂染色体的构象,可能会发现染色质组装的其它驱动力,并提供如基因组的区室化机制等折叠原理的见解。
2024年5月27日,深圳湾实验室分子生理学研究所的张浩岳课题组以及来自费城儿童医院的Gerd Blobel课题组在Nature Genetics 发表了题为 Genome folding principles uncovered in condensin-depleted mitotic chromosomes 的研究论文。该研究利用凝缩蛋白缺失的有丝分裂染色体,揭示了间期细胞中新型的基因组区室化机制。
作者首先利用CRISPR/Cas9基因编辑技术和AID快速蛋白降解系统标记了凝缩蛋白 I和II的共有亚基SMC2,从而构建了可靶向降解凝缩蛋白 I和II的细胞系。通过诺考达唑诱导的细胞周期停滞来富集前中期的细胞,然后进行植物生长素处理 0.5 小时、1 小时、4 小时或 8 小时,结果显示有丝分裂染色体的“杆状”形状逐渐消失,以及单个染色单体之间的广泛纠缠,证实了凝缩蛋白在维持有丝分裂染色体形态中的重要作用。
值得注意的是,在凝缩蛋白缺失后,有丝分裂染色体出现了区室化特征,并且区室化强度随着时间的推移逐渐加强和扩大。与之形成对应的是,拓扑关联结构域 (TAD) 和 CTCF/cohesin介导的结构性染色质环在这个过程中仍然无法被检测到。为了进一步揭示是哪种类型的凝缩蛋白缺失触发了有丝分裂的区室化。作者分别靶向标记了凝缩蛋白 I 的特异亚基 NCAPH 和凝缩蛋白 II 的特异亚基 NCAPH2,发现凝缩蛋白 I和凝缩蛋白 II均可以抑制有丝分裂区室化。
结合染色质组蛋白相关修饰特征和EV1聚类结果,作者鉴定了四种类型的有丝分裂期特有的染色质区室:mA1、mA2、mB1 和 mB4。其中;mA1、mA2属于常染色质区室;mB1 和 mB4属于异染色质区室。在分裂间期,mA1 和 mA2 的 EV1 值非常相似,使它们几乎无法区分;但在凝缩蛋白缺失的有丝分裂细胞中,mA1和mA2迅速分离。其中,mA1的EV1 值和同型相互作用(homotypic interactions)显着增加;而mA2 区室显示出较低的 EV1 值,并伴有减弱的同型相互作用。利用乙酰转移酶抑制剂A485预处理细胞后发现,有丝分裂期mA1区室化有部分减弱,提示H3K27ac 参与了凝缩蛋白缺失的有丝分裂细胞中部分 mA1 的区室化。mB1 区室代表以 H3K27me3 为标记的兼性异染色质;mB4区室代表H3K9me3为标记的组成型异染色质。在分裂间期细胞中,mB1 经常与组成型异染色质分离,显示阳性 EV1 值并被归类为常染色质 A 区室,而在有丝分裂期间发生反转与mB4共区室。mB4则在凝缩蛋白缺失的有丝分裂细胞中出现大量自聚集。
染色质区室化动力学的结果表明,mA1 在凝缩蛋白缺失后仅 30 分钟就开始出现,并在短短 1 小时内接近平台强度。相比之下,mB4 区室则随着时间的推移缓慢增强。mA1 和 mB4 区室形成速率的差异表明常染色质和组成型异染色质的区室化过程可以解偶联(如图所示)。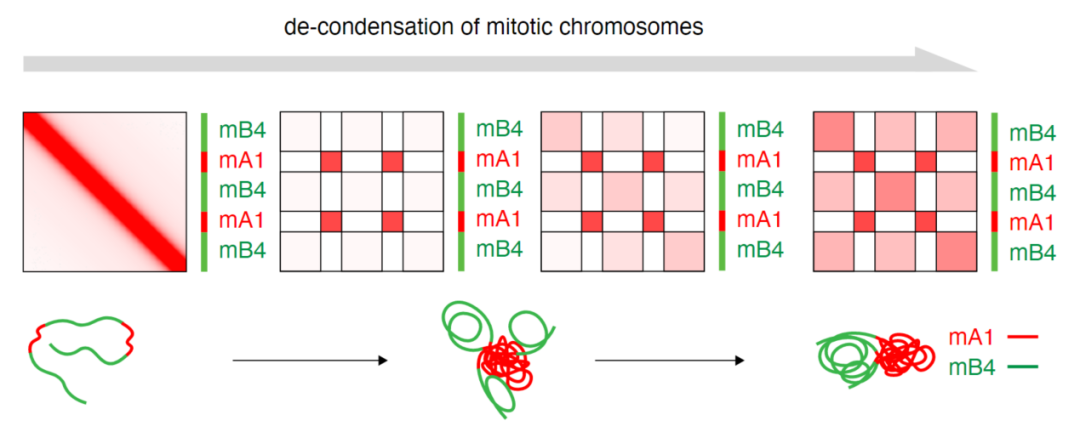
除了染色质区室化之外,作者还发现顺式作用元件(CRE,如启动子或者增强子)在凝缩蛋白降解30分钟后快速出现局部相互作用。这一结果令人感到非常惊讶,因为大部分可能介导CRE相互作用的染色质结合蛋白、转录因子、成环因子等在有丝分裂期均与会从染色质解离。这一现象说明CRE相互作用可能比人们预想中更加独立于这些蛋白的介导作用。另外,在凝缩蛋白降解后,转录并未恢复,因此单纯的CRE相互作用并不足以驱动转录活性。
有趣的是,作者利用ChIP-seq发现参与异染色质组装的HP1蛋白在有丝分裂期并不与染色质结合。因此,提示HP1蛋白可能不参与mB4组成性异染色质的区室化。为了验证这一可能,作者利用AID和dTAG快速蛋白降解系统构建了同时靶标HP1α, HP1β, HP1γ三种HP1蛋白的可诱导降解细胞系。值得注意的是作者发现在分裂期降解三种HP1蛋白并不会影响下一个细胞周期的组成性异染色质区室化,提示HP1蛋白家族并不参与异染色质区室化的形成。
综上,这项研究不仅揭示了在凝缩蛋白 I 和 II 共缺失后有丝分裂染色体复杂的区室化,还发现了基因组折叠的新原理,包括组成型异染色质自聚集能力和顺式调节元件接触的形成。这些发现为我们理解细胞周期中染色质的动态变化提供了新的视角。
深圳湾实验室分子生理学研究所特聘研究员张浩岳以及费城儿童医院Gerd Blobel教授为该文的通讯作者。张浩岳特聘研究员课题组研究助理赵韩为该文的第一作者。
原文链接:https://www.nature.com/articles/s41588-024-01759-x
参考文献
1.Nichols,M,H.& Corces,V.G. Principles of 3D compartmentalization of the humangenome. Cell reports 35,109330,doi:10.1016/j.celrep.2021.109330 (2021).
2.Zhang, H. et al. Chromatin structure dynamics during the mitosis-to-Gl phase transition. Nature 576,158-162,doi:10.1038/s41586-019-1778-y(2019).
3. Abramo,K.et al. A chromosome folding intermediate at the condensin-to-cohesin transition during telophase. Nature cell biology 21,1393-1402,doi:10.1038/s41556-019-0406-2(2019).
4. Pelham-Webb,B.et al. H3K27ac bookmarking promotes rapid post-mitotic activation ofthe pluripotent stem cell program without impacting 3D chromatin reorganization.Molecular cell 81,1732-1748 e1738, doi:10.1016/i.molcel.2021.02.032 (2021).
5. Zhang, H.& Blobel, G. A. Genome folding dynamics during the M-to-Gl-phase transitionCurrent opinion in genetics & development 80, 102036, doi:10.1016/.gde.2023.102036(2023).
6. Gibcus,y.H et al. A pathway for mitotic chromosome formation. Science 359doi:10.1126/science.aao6135 (2018).
7. Samejima, K. et al. Functional analysis after rapid degradation of condensins and 3D-EM reveals chromatin volume is uncoupled from chromosome architecture in mitosis. Journalofcell science 131, doi:10.1242/ics.210187(2018).